ŋšÔŗĘßfŧ°ûŖ¨APCsŖŠÖĐMHCˇÖ×ĶĩÄ×÷ĶÃCÖÆŧ°ßŨ
Ô ąëąë ÎÄÕÂíÔ´ŖēÉúÎīÖÆÆˇČĻ
Tŧ°ûŌĒąģŧ¤ģîŖŦËüĩÄĘÜķwąØí×ReĶÉĖØĘâ“ŋšÔŗĘßfŧ°û”ąíÃæĩÄMHCˇÖ×ĶŗĘŦFĩÄĩ°°×Ų|ÆŦļÎĄŖĶÉIîMHCˇÖ×ĶŗĘßfĩÄŋšÔŋÉŌÔ×ĸûĐÔTŧ°û“¸QŌ”ŧ°ûŖŦŌÔ´_ļ¨ËüĘĮˇņąģ¸ĐČž˛ĸĒÔąģ´Ũ§ĄŖĶÉIIîMHCˇÖ×ĶŗĘßfĩÄŋšÔĪōÃâŌßĪĩŊy°lŗöž¯ķŖŦĖáĐŅËü×ĸŌâ˛ģ¸ĐČžŧ°ûĩÄČëĮÖÕßŖŦ˛ĸÍÖú´_ąŖĸĶ´ķĩÄßmĒĐÔÃâŌßĪĩŊyĩÄQļ¨˛ģĘĮĶÉÎŧ°û×öŗöĩÄĄŖÔÚČËîČēķwÖĐŖŦĶĐÔSļāÂÔÎĸ˛ģÍŦĩÄMHCˇÖ×ĶĩÄģųŌōĄŖŌō´ËŖŦēÜŋÉÄÜÖÁÉŲĶĐŌģĐŠČËîūĶĐÄÜōÕšĘží×ÔČÎēβĄÔķwĩÄĩ°°×Ų|ÆŦļÎĩÄMHCˇÖ×ĶĄŖ

1.ŌũŅÔ
ÔÚËųĶĐŗÉÃâŌßĪĩŊyĩĸÅÄîÖĐŖŦŌ˛ÔS×îŅÅĄĸŽČģŌ˛×îŗöČËŌâÁĪĩÄĘĮŋšÔŗĘßfŖēŌģŧ°ûĪōÁíŌģŧ°ûŗĘŦFĩ°°×Ų|ÆŦļÎĩĸÅÄîĄŖÕũČįÄãĸŋ´ĩŊĩÄŖŦŋšÔŗĘßfĻßmĒĐÔÃâŌßĪĩŊyĩÄšĻÄÜÖÁęPÖØŌĒŖŦŗĘßfŋšÔŊoTŧ°ûĩÄŧ°û——ŋšÔŗĘßfŧ°ûŖ¨APCsŖŠ——°įŅŨÖøęPæIŊĮÉĢĄŖ×ÎŌĘ×ĪČĶÕAPCÉĪëHßMĐĐŗĘßfĩÄ“V¸æÅÆ”ŖēIîēÍIIîMHCˇÖ×ĶĄŖ
2.IîMHCˇÖ×Ķ
IîēÍIIîMHCˇÖ×ĶĩÄŊYŦFÔÚŌŅŊąģ×ĐŧˇÖÎöŖŦŌō´ËÃâŌßWŧŌĻß@ĐŠˇÖ×ĶĩÄÍâĶ^ĶĐÁËēÜēÃĩÄÁËŊâĄŖIîˇÖ×ĶĶĐŌģÉļˡâé]ĩÄŊYēĪ˛ÛŖŦŌō´ËËüŗĘŦFĩÄĐĄĩ°°×Ų|ÆŦļÎŖ¨ëÄŖŠąØíßmēĪŊYēĪ˛ÛĩÄžÖĪŪŋÕégŖ¨ČįšûÄãÔ¸ŌâŖŦŋÉŌÔˇQÖŽé“ĐĄÃæ°ü”ŖŠĄŖĘÂÉĪŖŦŽÃâŌßWŧŌÄIîˇÖ×ĶĩÄÕÆÎÕÖĐČĄŗöëIJĸĻÆäßMĐĐyĐōrŖŦËû°lŦF´ķļāĩëÄĩÄéLļČé8ģō9°ąģųËáĄŖß@ĐŠëÄÔÚÉļËąģå^ļ¨ŖŦéLļČĩÄÎĸĐĄ×ģ¯Í¨ß^ÔĘÔSëÄÔÚÖĐĐÄÉÔÎĸÍšÆđíßmĒĄŖ
ÃŋČËîļŧĶĐČũģųŌōŖ¨HLA-AĄĸHLA-BēÍHLA-CŖŠž´aIîMHCĩ°°×ŖŦÎģĶÚĩÚ6ĖČžÉĢķwÉĪĄŖŌōéÎŌĶĐÉĩÚ6ĖČžÉĢķwŖ¨Ōģí×ÔĸĶHŖŦŌģí×Ô¸¸ĶHŖŠŖŦÎŌÃŋČËŋš˛ĶĐÁųIîMHCģųŌōĄŖIîHLAĩ°°×ÅcÁíŌģˇNÃûéβ2-ÎĸĮōĩ°°×ĩÄĩ°°×ÅäĻŖŦŗÉÍęÕûĩÄIîMHCˇÖ×ĶĄŖÔÚČËîČēķwÖĐŖŦ´ķŧsĶĐ1500ˇNÂÔÎĸ˛ģÍŦĩÄģųŌōĐÎĘŊž´aČũˇNIîHLAĩ°°×ĄŖß@ĐŠHLA-AĄĸHLA-BēÍHLA-CģųŌō×ķwž´aĩÄĩ°°×´ķÖÂĐÎ îĪāÍŦŖŦĩĢËüÔÚŌģÉ°ąģųËáÉĪĶĐËų˛ģÍŦĄŖÃâŌßWŧŌĸžßĶĐļāˇNĐÎBĩġÖ×ĶˇQé“ļāBĐÔ”ŖŦIîHLAĩ°°×oŌɡûēĪß@ŌģÃčĘöĄŖĪāąČÖŽĪÂŖŦÎŌËųĶĐČËļŧĶĐĪāÍŦĩÄβ2-ÎĸĮōĩ°°×ģųŌōĄŖ
ĶÉĶÚËüĘĮļāBĐÔĩÄŖŦIîMHCˇÖ×ĶŋÉŌÔĶвģÍŦĩÄŊYēĪÄŖĘŊŖŦŌō´ËŋÉŌÔŗĘŦFÔÚÄŠļËžßĶвģÍŦîĐÍ°ąģųËáĩÄëÄĄŖĀũČįŖŦŌģĐŠIîMHCˇÖ×ĶÅcŌģļËžßĶĐĘčËŽĐÔ°ąģųËáĩÄëÄŊYēĪŖŦļøÆäËûMHCˇÖ×Ķt¸üĪ˛gÔÚß@Ōģå^ļ¨ÎģÖÞßĶĐAĐÔ°ąģųËáĄŖĶÉĶÚČËîĶĐŋÉÄÜąíß_ļāß_ÁųˇN˛ģÍŦĩÄIîˇÖ×ĶŖŦÎŌĩÄIîˇÖ×ĶŋķwÉĪŋÉŌÔŗĘŦFļāˇNļāĶĩÄëÄĄŖ´ËÍâŖŦąMšÜMHC IˇÖ×ĶĻëÄÉļËĩÄÄŗĐŠ°ąģųËáˇĮŗŖĖôĖŪŖŦĩĢËüÔÚĩ°°×Ų|ÆŦļÎÖĐĐÄĩÄ°ąģųËáßxņÉĪĪāŽëSŌâĄŖŌō´ËŖŦŌģŊoļ¨ĩÄIîMHCˇÖ×ĶŋÉŌÔŊYēĪ˛ĸŗĘŦFÔSļā˛ģÍŦĩÄëÄŖŦÃŋëÄļŧÅcÆäŊYēĪ˛ÛÄŠļË´æÔÚĩÄĖØļ¨°ąģųËá“ÆõēĪ”ĄŖ
3.IIîMHCˇÖ×Ķ
ĪņIîˇÖ×ĶŌģĶŖŦIIîMHCˇÖ×ĶŖ¨ĶÉĩÚ6ĖČžÉĢķwHLA-D ^ĶōĩÄģųŌōž´aŖŠŌ˛ĘĮOÆäļāBĐÔĩÄĄŖÔÚČËîČēķwÖĐŖŦ´ķŧsĶĐ700ˇN˛ģÍŦĩÄIIîMHCˇÖ×Ķ°æąžĄŖÅcIîMHCˇÖ×Ķ˛ģÍŦŖŦIIîMHCˇÖ×ĶĩÄŊYēĪ˛ÛÉļËļŧĘĮé_ˇÅĩÄŖŦŌō´ËëÄŋÉŌÔIJÛÖĐÉėŗöĄŖÕũČįÄãÄß@ŌģĖØÕ÷ÖĐŋÉÄÜîAÁĪĩŊĩÄŖŦÅcIIîˇÖ×ĶŊYēĪĩÄëÄąČÕŧūˇâé]˛ÛĩÄIîˇÖ×ĶĩÄëÄŌĒéL——ÔÚĘŽČũĩŊļūĘŽÎå°ąģųËáĩġļúČĄŖ´ËÍâŖŦĻĶÚIIîMHCˇÖ×ĶíÕfŖŦå^ļ¨ëÄĩÄęPæI°ąģųËáŅØÖøŊYēĪ˛ÛˇÖ˛ŧŖŦļø˛ģĘĮžÛŧ¯ÔÚÉļËĄŖ
4.IîMHCˇÖ×ĶĩÄŋšÔŗĘßf
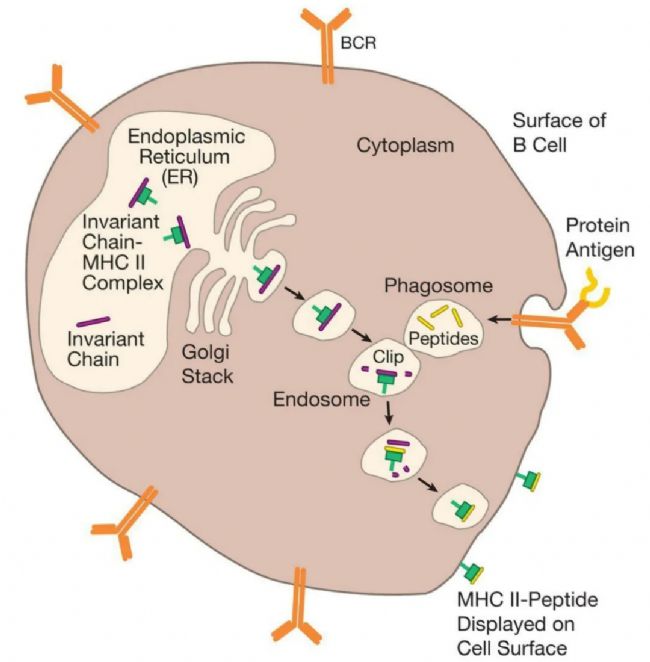
MHC IˇÖ×ĶĘĮŧ°ûąíÃæĩÄV¸æÅÆŖŦÕšĘžÔŧ°ûÖÆÔėĩÄĩ°°×Ų|ÆŦļÎĄŖÃâŌßWŧŌˇQß@ĐŠéČÔ´ĐÔĩ°°×ĄŖËü°üĀ¨ÆÕͨĩÄŧ°ûĩ°°×ŖŦČįøēÍŊYĩ°°×ŖŦŌÔŧ°ŋÉÄܸĐČžŧ°ûĩIJĄļžēÍÆäËûÎĸÉúÎīž´aĩÄĩ°°×ĄŖĀũČįŖŦŽ˛ĄļžßMČëŧ°ûrŖŦËüĀûĶÃŧ°ûÉúÎīēĪŗÉCÖÆŽaÉú˛ĄļžģųŌōž´aĩÄĩ°°×ĄŖČģēķŖŦß@ĐŠ˛Ąļžĩ°°×ĩÄĶąžÅcËųĶĐÕũŗŖŧ°ûĩ°°×ĩÄĶąžŌģÆđĶÉIîMHCˇÖ×ĶÕšĘžĄŖŌō´ËŖŦëHÉĪŖŦMHC IV¸æÅÆĐû÷ÁËŧ°ûČÕũÔÚÖÆÔėĩÄËųĶĐĩ°°×Ų|ĩÄĶąžĄŖ
ČËķw×ēõËųĶĐŧ°ûļŧÔÚÆäąíÃæąíß_IîˇÖ×ĶŖŦąMšÜˇÖ×ĶĩÄĩÁŋŌōŧ°ûļøŽĄŖĸûĐÔTŧ°ûŖ¨Ō˛ˇQéŧ°ûļžĐÔÁÜ°Íŧ°ûģōCTLsŖŠz˛éIîMHCˇÖ×ĶÕšĘžĩÄĩ°°×Ų|ÆŦļÎĄŖŌō´ËŖŦ×ēõËųĶĐŧ°ûļŧĘĮŌģąž“´ōé_ĩÄø”ŖŦŋÉŌÔĶÉCTLsz˛éŌÔ´_ļ¨ËüĘĮˇņąģ˛ĄÔķwČëĮÖ˛ĸĒÔąģ´Ũ§ĄŖŌģĩäĐÍĩÄČËîŧ°ûÔÚÆäąíÃæ´ķŧsĶĐ100,000IîˇÖ×ĶŖŦËüÔÚÄĮĀī´ķŧsŌģĖėēķŖŦMHCV¸æÅÆąģĐÂĩÄČĄ´ú——Ōō´ËIîMHCÕšĘžąŖŗÖ×îĐÂĄŖ
ČÔ´ĐÔĩ°°×ąģĖĀí˛ĸŅbŨdĩŊIîMHCˇÖ×ĶÉĪĩġŊĘŊˇĮŗŖĶĐȤĄŖŽmRNAÔÚŧ°ûĩÄŧ°ûŲ|ÖĐąģˇ×gŗÉĩ°°×rŖŦŊŗŖūˇ¸åeÕ`ĄŖß@ĐŠåeÕ`ŋÉÄܧÖÂÉúŽaŗö˛ģÕũ´_ÕÛ¯BĩÄoĶÃĩ°°×ĄŖ´ËÍâŖŦĩ°°×Ų|ŌōÕũŗŖÄĨpļøĘÜpĄŖŌō´ËŖŦéÁË´_ąŖÎŌĩÄŧ°û˛ģūŗäMĶĐČąĪŨĩÄĩ°°×ŖŦĶĐČąĪŨģōÄĨpĩÄĩ°°×ąģËÍČëŧ°ûŲ|ÖĐĩÄĩ°°×ÆÆÄ“CÆ÷”ÖĐŖŦß@ĐŠCÆ÷ĩÄšĻÄÜîËÆĶÚÄžĐŧCĄŖß@ĐŠĩ°°×ÄžĐŧCąģˇQéĩ°°×øķwŖŦËüĸĩ°°×Ų|ĮиîŗÉëÄĄŖČģēķŖŦß@ĐŠëÄÖĐĩÄ´ķļāĩąģßMŌģ˛ŊˇÖŊâŗÉΰąģųËáŖŦß@ĐŠ°ąģųËáąģÖØĐÂĶÃíÖÆÔėĐÂĩ°°×ĄŖČģļøŖŦĶÉĩ°°×øķwÔėĩÄŌģĐŠëÄĶÉĖØļ¨ĩÄŪDß\ĩ°°×Ŗ¨TAP1ēÍTAP2ŖŠ´Šß^ĤßMČëČŲ|žWŖ¨ERŖŠ——ß@ĘĮŌģ´ķĩÄĄĸÄŌ îĩÄŊYŖŦ´ķļāĩ×ĸļ¨ŌĒß\ŨĩŊŧ°ûąíÃæĩÄĩ°°×Äß@Āīé_ĘŧËüĩÄÂÃŗĖĄŖ
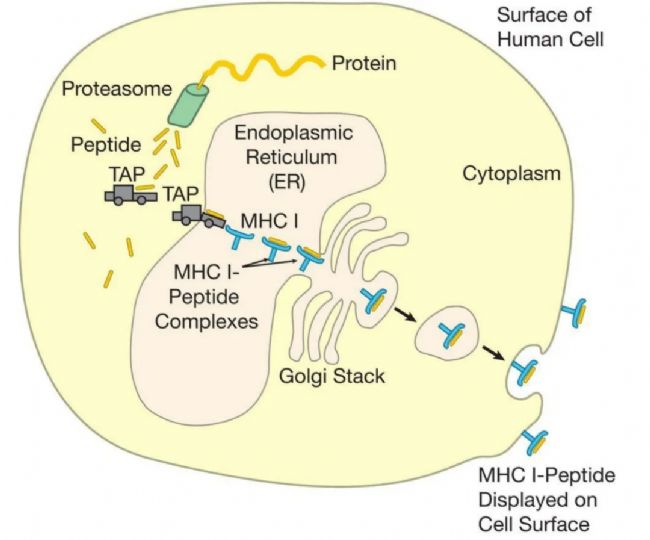
ŌģĩŠßMČëČŲ|žWŖŦŌģĐŠëÄļÎąģßxÖĐŅbŨdĩŊIîMHCˇÖ×ĶĩÄ°ŧ˛ÛÖĐĄŖÎŌÕf“ßxÖĐ”ŖŦĘĮŌōéŖŦÕũČįÎŌĶÕĩÄŖŦ˛ģĘĮËųĶĐĩÄëÄļÎļŧÄÜßmÅäĄŖĘ×ĪČŖŦŌģëÄļÎąØíĘĮÕũ´_ĩÄéLļČ——´ķŧsžÅ°ąģųËáĄŖ´ËÍâŖŦëÄļÎÉļËĩÄ°ąģųËáąØíÅcMHCˇÖ×Ķ°ŧ˛ÛÉļËĩÄå^ļ¨°ąģųËáĪāČŨĄŖī@ČģŖŦĶÉĩ°°×øķwĘäĩÄ“ģI´a”˛ĸˇĮļŧžßĶĐß@ĐŠĖØÕ÷ŖŦÄĮĐŠ˛ģˇûēĪĩÄĸąģŊĩŊâģōß\ģØČŲ|žWÍâĩÄŧ°ûŲ|ÖĐĄŖŌģĩŠIîMHCˇÖ×ĶŅbŨdÁËëÄļÎŖŦËüžÍūĮ°Íųŧ°ûąíÃæßMĐĐÕšĘžĄŖŌō´ËŖŦĘäIîÕšĘžĶĐČũÖ÷ŌĒ˛ŊķEŖēĩ°°×øķwÉúŗÉëÄļÎŖŦTAPŪDß\ĩ°°×ĸëÄļÎß\ËÍĩŊČŲ|žWŖŦŌÔŧ°ëÄļÎÅcMHC IˇÖ×ĶĩÄ°ŧ˛ÛŊYēĪĄŖ
ÔÚÆÕͨŧ°ûÖĐŖŦČį¸Îŧ°ûēÍĐÄŧĄŧ°ûŖŦĩ°°×øķwĩÄÖ÷ŌĒšĻÄÜĘĮĖĀíĶĐČąĪŨĩÄĩ°°×Ų|ĄŖŌō´ËŖŦÄãŋÉŌÔĪëĪķŖŦß@ĐŠŧ°ûÖĐĩÄ“ĮиîÆ÷”˛ĸ˛ģĖĢÔÚŌâĩ°°×Ų|ĘĮČįēÎąģĮиîĩÄ——ËüÖģĘĮëSŌâĮиîĄŖŊYšûŖŦŌģĐŠëÄļÎĸßmēĪMHCÕšĘžŖŦĩĢ´ķļāĩ˛ģūĄŖĪāąČÖŽĪÂŖŦÔÚŖéTÕšĘžŋšÔĩÄŧ°ûŖŦČįžŪĘÉŧ°ûÖĐŖŦß@ˇNĮиî˛ĸ˛ģÄĮôëSCĄŖĀũČįŖŦIFN-γÅcžŪĘÉŧ°ûąíÃæĩÄĘÜķwŊYēĪÉĪÕ{ÁËČũˇNÃûéLMP2ĄĸLMP7ēÍMECL1ĩÄĩ°°×Ų|ĩÄąíß_ĄŖß@ĐŠĩ°°×Ų|ČĄ´úÁËÕũŗŖĩ°°×øķwCĐĩÖĐĩÄČũ“ė´æ”ĩ°°×Ų|ĄŖß@ˇNĖæQĩÄŊYšûĘĮŖŦ“ļ¨ÖÆ”ĩÄĩ°°×øķwŦFÔÚ¸üAĪōĶÚÔÚĘčËŽģōAĐÔ°ąģųËáēķĮиîĩ°°×Ų|ĄŖÄãéĘ˛Ã´ŖŋŌōéTAPŪDß\ĩ°°×ēÍMHC IˇÖ×ĶļŧÆĢÛžßĶĐĘčËŽģōAĐÔCÄŠļËĩÄëÄļÎĄŖŌō´ËŖŦÔÚŋšÔŗĘßfŧ°ûÖĐŖŦËĘĩ°°×øķwąģĐŪ¸ÄŖŦŌÔąãËüÄÜŽaÉúļ¨ÖÆĩÄëÄļÎŖŦÄļøĖá¸ßIîÕšĘžĩÄЧÂĘĄŖ
ĩ°°×øķwĻËüŽaÉúĩÄëÄļδķĐĄŌ˛˛ģĖĢÔÚŌâŖŦŧČČģIîÕšĘžĩÄÉņÆæĩ×Ö´ķŧsĘĮžÅ°ąģųËáŖŦÄãŋÉÄÜūĪëĪķČŲ|žWĸŗäŗâÖøoĶÃĩÄëÄļÎŖŦËüŌĒôĖĢéLŌĒôĖĢļĖĄŖČģļøŖŦŊYšûąíÃ÷TAPŪDß\ĩ°°×Ļ°ËĩŊĘŽÁų°ąģųËáéLĩÄëÄļΞßĶĐ×î¸ßĩÄĶHēÍÁĻĄŖŌō´ËŖŦTAPŪDß\ĩ°°×ēYßxĩ°°×øķwŽaÉúĩÄëÄļÎŖŦ˛ĸĪČß\ŨÄĮĐŠžßĶĐÕũ´_CÄŠļˡNîĮŌéLļČ´ķÖÂÕũ´_ĩÄëÄļÎĄŖŌģĩŠēōßxëÄļÎąģß\ËÍĩŊČŲ|žWŖŦøūĐŪŧôĩôļāĶāĩÄNÄŠļË°ąģųËáŖŦĘšëÄļδķĐĄßmēĪÅcIîMHCˇÖ×ĶŊYēĪĄŖ
ß@“ĮĐËé˛ĸÕšĘž”ĪĩŊyĩÄŌģÖØŌĒĖØücĘĮŖŦĩ°°×øķwĮĐËéĩÄ´ķļāĩĩ°°×Ų|ĘĮŊYÉĪĶĐČąĪŨĩÄĐÂēĪŗÉĩ°°×Ų|——ļø˛ģĘĮĐčŌĒģØĘÕĩÄÅfĩÄĄĸÄĨpĩÄĩ°°×Ų|ĄŖŌō´ËŖŦIîMHCˇÖ×ĶÕšĘžĩÄ´ķļāĩëÄļÎÔ´×ÔĐÂēĪŗÉĩÄĩ°°×Ų|ŖŦß@ĘšĩÃÃâŌßĪĩŊyÄÜōŋėËŲĻ¸ĐČž×öŗöˇ´ĒĄŖ
5.IIîMHCˇÖ×ĶĩÄŋšÔŗĘßf
ÅcIîMHCˇÖ×ĶÖŧÔÚĪōĸûĐÔTŧ°ûÕšĘžĩ°°×Ų|ÆŦļβģÍŦŖŦIIîMHCˇÖ×ĶĪōŨoÖúTŧ°ûÕšĘžëÄļÎĄŖÅc×ēõÔÚÃŋˇNŧ°ûÉĪļŧąíß_ĩÄIîMHCˇÖ×Ķ˛ģÍŦŖŦIIîˇÖ×ĶHÔÚÃâŌßĪĩŊyĩÄŧ°ûÉĪąíß_ĄŖß@ĘĮĶĐĩĀĀíĩÄĄŖIîˇÖ×ĶŖéTÕšĘžÔÚŧ°ûČÖÆÔėĩÄĩ°°×Ų|ŖŦŌō´ËIîˇÖ×ĶĩÄÆÕąéĐÔéCTLĖᚊÁËz˛éķwČ´ķļāĩŧ°ûĘĮˇņ¸ĐČžĩÄCūĄŖÁíŌģˇŊÃæŖŦIIîMHCˇÖ×ĶĩÄšĻÄÜžÍĪņV¸æÅÆŖŦĐû÷ŧ°ûÍâ°lÉúĩÄĘÂĮéŖŦĖáĐŅŨoÖúTŧ°û×ĸŌâÎŖëUĄŖŌō´ËŖŦĖĐĐß@íČÎÕËųĐčĩÄąíß_IIîĩÄŧ°ûĪāĻŨ^ÉŲ——ÖģŌĒ×ãōÔÚÉíķwĩĸ÷˛ŋÎģ˛ÉĶhžŗŧ´ŋÉĄŖ
ŗÉIIîMHCˇÖ×ĶĩÄɡNĩ°°×Ų|Ŗ¨ˇQéαēÍβæŖŠÔÚŧ°ûŲ|ÖĐŽaÉúŖŦ˛ĸąģ×ĸČëČŲ|žWŖŦÔÚÄĮĀīËüÅcŌģˇNÃûé˛ģ×æĩÄĩÚČũĩ°°×Ų|ŊYēĪĄŖß@ˇN˛ģ×æĩ°°×ĖĐĐךĻÄÜĄŖĘ×ĪČŖŦËüÎģĶÚMHC IIˇÖ×ĶĩÄ°ŧ˛ÛÖĐŖŦˇĀÖšËüÔÚČŲ|žWÖĐėÆđÆäËûëÄļÎĄŖß@ēÜÖØŌĒŖŦŌōéČŲ|žWŗäMÁËĶÉĩ°°×øķwĖĀíĩÄČÔ´ĐÔëÄļÎŖŦß@ĐŠëÄļÎąģŅbŨdĩŊIîMHCˇÖ×ĶÉĪĄŖČįšûß@ĐŠĩ°°×Ų|ÆŦļÎąģŅbŨdĩŊIIîˇÖ×ĶÉĪŖŦÄĮôIîēÍIIîMHCˇÖ×ĶĸÕšĘžĪāÍŦîĐÍĩÄëÄļÎŖēÄĮĐŠĶÉŧ°ûČŽaÉúĩÄĩ°°×Ų|ÖÆŗÉĩÄĄŖĶÉĶÚÄŋËĘĮ×IIîMHCˇÖ×ĶÕšĘží×Ôŧ°ûÍâĩÄĩ°°×Ų|ÆŦļÎŖŦŧ´ÍâÔ´ĐÔëÄļÎŖŦ˛ģ×æĖĐĐÁËŌģÖØŌĒšĻÄÜŖēËüŗäŽŌģ“°éÄī”ŖŦ´_ąŖ“˛ģßmŽĩÄסĮķÕß”Ŗ¨ČÔ´ĐÔëÄļÎŖŠ˛ģūąģMHC IIˇÖ×ĶÔÚČŲ|žWÖĐėÆđĄŖ
˛ģ×æĩÄÁíŌģšĻÄÜĘĮŌũ§IIîMHCˇÖ×Ķͨß^¸ß ģųļŅŖĩŊŧ°ûŲ|ÖСQéČķwĩÄĖØĘâÄŌÅŨĄŖÕũĘĮÔÚČķwÖĐŖŦIIîMHCˇÖ×ĶŅbŨdëÄļÎĄŖÄŋĮ°ĩÄËŧŋŧĘĮŖŦŽIIîMHCˇÖ×ĶÄČŲ|žWĪōČķwĐĐßMrŖŦŧ°ûÍâŌėĩÄĩ°°×Ų|ąģ°üēŦÔÚŌģÍĖĘÉķwÖĐŖŦ˛ĸąģ§Čëŧ°ûĄŖČģēķß@ÍĖĘÉķwÅcČķwēĪ˛ĸŖŦČķwÖĐĩÄøĸÍâÔ´ĐÔĩ°°×Ų|ĮĐËéŗÉëÄļÎĄŖÔÚ´ËÆÚégŖŦČķwøŌ˛ÆÆÄÁ˲ģ×æĩÄËųĶвŋˇÖŖŦŗũÁËëHÉĪĘØĐlMHCˇÖ×Ķ°ŧ˛ÛĩÄCLIPÆŦļÎĄŖÁîČËķ@Ķ ĩÄĘĮŖŦąMšÜÍâÔ´ĐÔĩ°°×Ų|ēͲģ×æąģČķwÖĐĩÄøĮĐËéŖŦIIîMHCˇÖ×ĶąžÉí sēÁ°lopĄŖß@ŋÉÄÜĘĮŌōéMHCˇÖ×ĶĮÉÃîĩØÕÛ¯BŖŦĘšÃ¸oˇ¨ŊĶĶ|ĩŊËü×îĪ˛gĩÄĮиîÎģücĄŖ
Åc´ËÍŦrŖŦŌģˇNŧ°ûĩ°°×HLA-DMŌ˛ĩŊß_ÁËČķwŖŦ´ßģ¯“ÕŧÎģˇû”CLIPĩÄáˇÅĄŖß@ÔĘÔSŌģÍâÔ´ĐÔëÄļÎąģŅbŨdĩŊŦFÔÚŌŅŊŋÕĩÄIIîMHCˇÖ×ĶĩÄ°ŧ˛ÛÖĐĄŖĩĢHLA-DM˛ģHHĘĮĖßŗöCLIPéëÄļÎōvŗöŋÕégĄŖHLA-DMÅcÔÚëÄļθ ÅcIIîMHCˇÖ×ĶĩÄŊYēĪŖŦ´_ąŖÖģĶĐžoÃÜŊYēĪĩÄëÄļβÅūąģÕšĘžĄŖ×îēķŖŦMHCŧĶëÄļÎÍēĪÎīąģß\ËÍĩŊŧ°ûąíÃæßMĐĐÕšĘžĄŖ

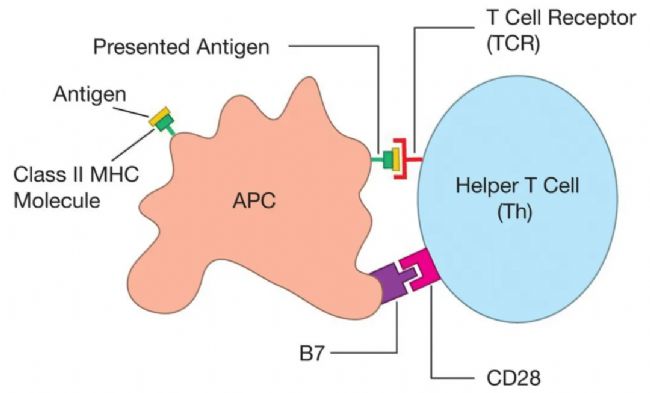
ÖØŌĒĩÄĘĮŌĒÕJ×RĩŊIîēÍIIîMHCˇÖ×ĶĶĐÉĒÁĸĩÄŅbŨdÎģücēÍÍžŊĄŖÕũĘĮß@ˇNŅbŨdÎģücēÍÍžŊĩġÖëxŖŦĘšĩÃIîV¸æÅÆÄÜōĐû÷ŧ°ûȲŋ°lÉúĩÄĘÂĮéŖ¨ŊoĸûĐÔTŧ°ûŖŠŖŦļøIIîV¸æÅÆÄÜōĐû÷ŧ°ûÍâ˛ŋ°lÉúĩÄĘÂĮéŖ¨ŊoŨoÖúTŧ°ûŖŠĄŖ
6.ŋšÔŗĘßfŧ°û
ÔÚĸûĐÔTŧ°ûÄÜōĸËĀÄŋËģōŨoÖúTŧ°ûÄÜōĖᚊÍÖúÖŽĮ°ŖŦËüąØíąģŧ¤ģîĄŖé´ËŖŦŌģTŧ°ûąØí×ReĶÉÁíŌģŧ°ûąíÃæĩÄMHCˇÖ×ĶŗĘßfĩÄĪāĒŋšÔĄŖĩĢß@߲ģōĄŖËüßąØíŊĶĘÕĩŊĩÚļūš˛´Ėŧ¤ĐÅĖĄŖÖģĶĐÄŗĐŠŧ°ûŅbäÁËĖᚊIîēÍIIîMHCÕšĘžŌÔŧ°š˛´Ėŧ¤ĩÄÄÜÁĻĄŖß@ĐŠžÍĘĮŋšÔŗĘßfŧ°ûŖ¨APCsŖŠĄŖ
ĶÉĶÚŋšÔŗĘßfŧ°ûĩÄš¤×÷ĘĮŧ¤ģîĸûĐÔēÍŨoÖúĐÔTŧ°ûŖŦß@ĐŠŧ°ûąžĒÔąģÃüÃûé“Tŧ°ûŧ¤ģîŧ°û”ĄŖß@ĸąÜÃâÁËÅcķwČÆÕͨŧ°ûĩÄģėĪũŖŦß@ĐŠÆÕͨŧ°û˛ģÄÜŧ¤ģîTŧ°ûŖŦĩĢËü´_ĘšĶÃIîMHCˇÖ×ĶíŗĘßfß@ĐŠŧ°ûČŽaÉúĩÄŋšÔŖŦŌÔĖáĐŅĸûĐÔTŧ°ûĄŖÄãĶXĩÃÃâŌßWŧŌĘĮ˛ģĘĮžÍĪ˛g°ŅĘÂĮéÅĒĩÃÍësŖŋÎŌĶĐrß@ôÕJéĄŖoÕČįēÎŖŦéÁËąŖŗÖĮåÎúŖŦÕĶץŖŦ“ŋšÔŗĘßfŧ°û”ß@ĐgÕZŋĘĮÖ¸ÄĮĐŠÄÜōĖᚊTŧ°ûŧ¤ģîËųĐčĩĸßËŽÆŊMHCĩ°°×ēÍš˛´Ėŧ¤ˇÖ×ĶĩÄĖØĘâŧ°ûĄŖ
š˛´Ėŧ¤Í¨ŗŖÉæŧ°ŋšÔŗĘßfŧ°ûąíÃæĩÄŌģÃûéB7ĩÄĩ°°×Ų|ŖŦËü“˛åČë”Tŧ°ûąíÃæĩÄŌģÃûéCD28ĩÄĩ°°×Ų|ĄŖ
ŌŅŊ´_ļ¨ÁËČũˇNîĐÍĩÄŋšÔŗĘßfŧ°ûŖēģîģ¯ĩÄäÍģ îŧ°ûĄĸģîģ¯ĩÄžŪĘÉŧ°ûēÍģîģ¯ĩÄBŧ°ûĄŖËųĶĐß@ĐŠļŧŲĶÚ°×ŧ°ûŖŦĶÉĶÚĐÂĩÄŅĒŧ°ûū˛ģāŽaÉúŖŦŌō´ËŋšÔŗĘßfŧ°ûŖ¨APCsŖŠŋÉŌÔ¸ųūĐčŌĒĩÃĩŊŅaŗäĄŖ
6.1.ģîģ¯ĩÄäÍģ îŧ°û
äÍģ îŧ°ûŖ¨DCsŖŠžßĶĐĖØÕ÷ĐÔĩÄĐĮĐÎĐÎ îŖŦÆäÃûˇQíÔ´ĶÚ“äÍģ”ŌģÔ~ŖŦÔÔ~ͨŗŖĶÃĶÚÃčĘöÉņŊŧ°ûÉĪĩÄÍģÆđĄŖĐčŌĒ×ĸŌâĩÄĘĮŖŦß@ĐŠŧ°ûÅcÖŽĮ°ĖáĩŊĩÄ{ŧ°ûĶäÍģ îŧ°ûŖ¨pDCsŖŠˇĮŗŖ˛ģÍŦ——pDCsĩÄÖ÷ŌĒšĻÄÜĘĮÔÚ˛Ąļž¸ĐČžrŽaÉú´ķÁŋĩĸÉ_ËØαēÍβĄŖëHÉĪŖŦąMšÜËüąģˇQé“äÍģ îŧ°û”ŖŦėoÖšĩÄpDCsĪņBŧ°ûēÍTŧ°ûŌģĶŗĘAĐÎĄŖÖģĶĐÔÚËüąģ˛Ąļž¸ĐČžŧ¤ģîēķŖŦ˛ÅūŗĘŦFŗöäÍģ îŧ°ûĩÄĐĮĐÎĄŖäÍģ îŧ°ûÔøąģÕJéĘĮŌģˇNĪĄÆæŦFĪķĄŖČģļøŖŦŦFÔÚČËÕJ×RĩŊß@ĐŠŧ°ûĘĮËųĶĐŋšÔŗĘßfŧ°ûÖĐ×îÖØŌĒĩÄ——ŌōéäÍģ îŧ°ûŋÉŌÔͨß^ŧ¤ģîÔĘŧTŧ°ûíĸĶÃâŌߡ´ĒĄŖŌÔĪÂĘĮÆ䚤×÷ÔĀíĄŖ
×îŗõÃčĘöĩÄäÍģ îŧ°ûĘĮĐĮĐÎĩÄ“ĀʸņhËš”ŧ°ûŖŦËüÎģĶÚƤÄwĪ¡ŊĩÄŊMŋÖĐĄŖČģļøŖŦ´ËēķÔÚČĢÉí¸÷Ėļŧ°lŦFÁËäÍģ îŧ°ûĄŖŦFÔÚĮåŗūĩÄĘĮŖŦß@ĐŠäÍģ îŧ°ûĘĮÉÚąøŧ°ûŖŦËüÎģĶÚ´úąíÎŌĩÚŌģĩĀˇĀžĩÄÉĪƤŧ°ûÆÁÕĪÖŽĪÂĄŖÔÚÕũŗŖŊMŋŖ¨Î´¸ĐČžĩÄŊMŋŖŠÖĐŖŦDCsîËÆĶÚÆˇžÆĄŖąMšÜËüÃŋĐĄrŋÉŌÔÎüĘÕ´ķŧsËÄąļĶÚ×ÔÉíķwˇeĩÄŧ°ûÍâŌēŖŦĩĢËü´ķļāÖģĘĮÎüĘÕēķÔŲÅÅŗöĄŖÔÚß@ˇN“ėoÖš” îBĪÂŖŦDCsąíß_ŌģĐŠB7ēÍĪāĻŨ^ĩÍËŽÆŊĩÄÖ÷ŌĒŊMŋĪāČŨĐÔÍēĪķwŖ¨MHCŖŠˇÖ×ĶÔÚÆäąíÃæĄŖŌō´ËŖŦËü˛ģÉÃéLĪōTŧ°ûŗĘßfŋšÔŖŦĶČÆäĘĮĪōÔĘŧTŧ°ûŗĘßfĄŖß@ĘĮŌōéÔĘŧTŧ°ûĐčŌĒͨß^MHC-ëÄÍēĪÎīßMĐĐVˇēĩÄĘÜķwŊģÂŌÔŧ°´ķĩÄš˛´Ėŧ¤˛ÅÄÜąģŧ¤ģîĄŖ
Čįšû°lÉúÎĸÉúÎīČëĮÖŖŦäÍģ îŧ°ûËųÔÚĩÄŊMŋ×ŗÉÁËđöŖŦäÍģ îŧ°ûĸ×ĩÓģîģ¯”ĄŖDCsŋÉŌÔͨß^í×Ô ĸÅcđļˇĩÄÆäËûÃâŌßĪĩŊyŧ°ûĩÄĐÅĖąģŧ¤ģîĄŖĀũČįŖŦŽÖĐĐÔÁŖŧ°ûēÍžŪĘÉŧ°ûÔD´Ũ§ČëĮÖÕßrŖŦËüūáˇÅÄ[ÁöÄËĀŌō×ĶŖ¨TNFŖŠŖŦß@ˇNđļˇŧ°ûŌō×ĶŋÉŌÔŧ¤ģîDCsĄŖ´ËÍâŖŦ˛Ąļž¸ĐČžĩÄŧ°ûáˇÅĩĸÉ_ËØαģōβŌ˛ŋÉŌÔĶ|°lDCĩÄŧ¤ģîĄŖ×îēķŖŦäÍģ îŧ°ûžßĶĐÄŖĘŊ×ReĘÜķwŖ¨ĀũČįŖŦTollĶĘÜķwŖŠŖŦËüĶÃß@ĐŠĘÜķwí×ReČëĮÖÕßVˇēîeĩÄĖØÕ÷ĐÔˇÖ×ĶÄŖĘŊĄŖß@ĐŠÄŖĘŊ×ReĘÜķwŊĶĘÕĩŊĩÄĐÅĖÔÚŧ¤ģîäÍģ îŧ°ûÖĐŋÉŌÔ°l]ÖØŌĒ×÷ĶÃĄŖ
6.1.1.äÍģ îŧ°ûĩÄßwŌÆ
ŽäÍģ îŧ°ûąģđļˇŧ°ûŌō×Ķŧ¤ģîŖŦąģ´šËĀŧ°ûáˇÅĩÄģ¯WÎīŲ|ŧ¤ģîŖŦąģÆäÄŖĘŊ×ReĘÜķwĩÄŊYēĪŧ¤ģîŖŦģōĶÉß@ĐŠĐÅĖĩÄŊMēĪŧ¤ģîrŖŦ“ÆˇžÆ”ĩÄÉúģîˇŊĘŊū°lÉúžŪ´ķ×ģ¯ĄŖäÍģ îŧ°û˛ģÔŲ“ÆˇLēÍÍÂŗö”ĄŖŦFÔÚËü“ÍĖĪ”ÁËËüËųÎüĘÕĩÄ|Î÷ĄŖͨŗŖŖŦŌģDCÔÚŧ¤ģîēķÔÚŊMŋÖĐÍŖÁô´ķŧsÁųĐĄrŖŦĘÕŧ¯đļˇŋšÔĩÄ´úąíĐÔĶąžĄŖÄĮrŖŦÍĖĘÉ×÷ĶÃÍŖÖšŖŦģîģ¯ĩÄäÍģ îŧ°ûëxé_ŊMŋŖŦͨß^ÁÜ°ÍĪĩŊyĮ°Íų×îŊüĩÄÁÜ°ÍŊYĄŖÕũĘĮÆäÔÚŧ¤ģîrĩÄßwŌÆÄÜÁĻĘšĩÃäÍģ îŋšÔŗĘßfŧ°ûČį´ËĖØeĄŖ
ÔÚėoÖšĩÄDCȲŋĶĐ´ķÁŋĩÄ“Ļä”IIîMHCˇÖ×ĶĄŖŽŌģėoÖšĩÄDCąģŧ¤ģî˛ĸé_Ęŧ“ŗÉĘė”rŖŦß@ĐŠIIîMHCˇÖ×Ķé_ĘŧąģđöĩÄŋšÔŅbŨdĄŖĩŊDCĩŊß_ÄŋĩÄĩØ——ͨŗŖĐčŌĒ´ķŧsŌģĖėĩÄrég——ß@ĐŠŅbŨdÁËđļˇŋšÔĩÄIIîMHCˇÖ×ĶĸÔÚŧ°ûąíÃæī@ÖøÕšĘžĄŖÔÚÂÃÍžÖĐŖŦDCßÉĪÕ{ÆäIîMHCˇÖ×ĶĩÄąíß_ĄŖŌō´ËŖŦČįšûäÍģ îŧ°ûÔÚđöąģ˛Ąļž¸ĐČžŖŦĩŊËüĩŊß_ÁÜ°ÍŊYrŖŦ˛Ąļžĩ°°×ÆŦļÎĸÕšĘžÔÚäÍģ îŧ°ûĩÄIîMHCV¸æÅÆÉĪĄŖ×îēķŖŦÔÚßwŌÆß^ŗĖÖĐŖŦDCÔöŧĶÁËB7š˛´Ėŧ¤ĩ°°×ĩÄŽaÉúĄŖËųŌÔŽËüĩŊß_ÁÜ°ÍŊYrŖŦŗÉĘėĩÄäÍģ îŧ°ûíĶĐŧ¤ģîÔĘŧTŧ°ûËųĐčĩÄŌģĮĐŖē¸ßËŽÆŊĩÄIîēÍIIîMHCˇÖ×ĶŅbŨdÖøßmŽĩÄëÄļÎŖŦŌÔŧ°´ķÁŋĩÄB7ĩ°°×ĄŖ
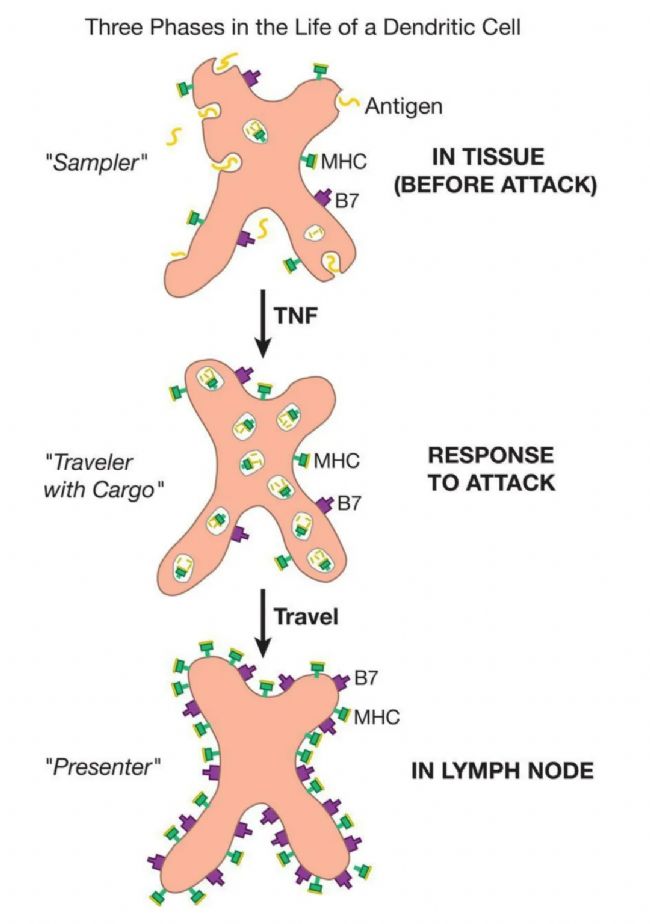
ŦFÔÚŖŦéĘ˛Ã´ÄãÕJéäÍģ îŧ°ûŖ¨DCsŖŠÔÚŊMŋÖĐVˇē˛ÉĶŋšÔēķŖŦÔÚé_ĘŧĮ°ÍųÁÜ°ÍŊYĩÄÂÃŗĖrūÍŖÖš˛ÉĶÄØŖŋŽČģĄŖäÍģ îŧ°ûĻ“Į°ž”°lÉúĩÄĮérČĄŌģ“ŋėÕÕ”ŖŦ˛ĸĸß@DĪņ§ĩŊÁÜ°ÍŊY——ÄĮĀīĘĮδŧ¤ģîĩÄTŧ°ûžÛŧ¯ĩÄĩØˇŊĄŖÔÚÄĮĀīŖŦÂÃĐĐÖĐĩÄäÍģ îŧ°ûŧ¤ģîÄĮĐŠTŧ°ûĘÜķwÄÜō×Re“DÆŦ”ÖĐĩÄČëĮÖÕßĩÄδŧ¤ģîTŧ°ûĄŖđļˇŧ°ûŌō×ĶČįÄ[ÁöÄËĀŌō×ĶŖ¨TNFŖŠĶ|°lDCsßwŌÆĩŊÁÜ°ÍŊYĩÄĘÂŌ˛ÍęČĢēĪĀíĄŖŽ žšŖŦÄãĪŖÍûäÍģ îŧ°ûŗÉĘėŖŦßwŌÆĩŊÁÜ°ÍŊYŖŦ˛ĸÖģÔÚđļˇßMĐĐrŗĘŦFŋšÔĄŖ
ŌģĩŠäÍģ îŧ°ûĩŊß_ÁÜ°ÍŊYŖŦËü´ķŧsÖģÄÜ´æģîŌģÖÜĄŖß@ˇNļĖēĩÄÛÃüÕ§Ōģŋ´ŋÉÄÜī@ĩÃÆæšÖĄŖŽ žšŖŦß@˛ĸ]ĶĐŊoäÍģ îŧ°ûēÜéLĩÄrégÅc“Õũ´_”ĩÄδŧ¤ģîTŧ°ûĪāĶöŖŦēķÕßÕũÔÚÁÜ°ÍŊYÖĐŅhŖŦ¤ÕŌÆäĪāĒĩÄŋšÔĄŖČģļøŖŦäÍģ îŧ°ûÃŋĐĄrŋÉŌÔÅcĩ°ŲÉõÖÁĩĮ§Tŧ°ûĪāģĨ×÷ĶÃŖŦËüļĖēĩÄŗĘŦFÛÃü´_ąŖÁËäÍģ îŧ°ûy§ĩÄĘĮ×îĐÂĩÄđļˇŋėÕÕĄŖ´ËÍâŖŦÔÚäÍģ îŧ°ûąģŧ¤ģîĩĢÔÚé_ĘŧÂÃĐĐÖŽĮ°ŖŦËüūŽaÉúĖØĘâĩÄŧ°ûŌō×ĶŖ¨Ú ģ¯Ōō×ĶŖŠŖŦß@ĐŠŌō×ĶšÄîˇQéÎēËŧ°ûĩÄ°×ŅĒŧ°ûëxé_ŅĒŌēŖŦßMČëŊMŋŖŦ˛ĸ×ŗÉäÍģ îŧ°ûĄŖŌō´ËŖŦŧ¤ģîĩÄäÍģ îŧ°ûÕĐÄŧ×ÔŧēĩÄĖæ´úÆˇĄŖß@ĐŠĐÂÕĐÄŧĩÄDCsŋÉŌÔĸđļˇĩÄ×îĐÂDĪņ§ĩŊÁÜ°ÍŊYŖŦŌōéđļˇßÔÚĀ^ĀmĄŖ
äÍģ îŧ°ûļĖēÛÃüĩÄÁíŌģÔŌōĘĮÎŌÔÚĩÚļūÖvÖĐĖáĩŊĩÄŖŦÃâŌߡ´ĒĩÄļČÅcšĨôĩÄĀÖØŗĖļČŗÉąČĀũĘĮˇĮŗŖÖØŌĒĩÄĄŖäÍģ îŧ°ûĩÄļĖēÛÃüĶĐÖúĶÚŦFß@ŌģücĄŖŌÔĪÂĘĮČįēÎŦFĩÄĄŖÔÚÎĸÉúÎīšĨôÆÚégŖŦŧ¤ģîĩÄTŧ°ûĩÁŋĸČĄQĶÚĸđļˇĪûĪĸ§ĩŊ¸ŊŊüÁÜ°ÍŊYĩÄŗÉĘėäÍģ îŧ°ûĩÄĩÁŋĄŖČįšûšĨôŨ^ČõŖŦđļˇÖĐĩÄžŪĘÉŧ°ûŽaÉúĩÄŧ°ûŌō×ĶĪāĻŨ^ÉŲŖŦÖģĶĐÉŲĩäÍģ îŧ°ûy§ËüĩÄØÎīąģÅÉĮ˛ĄŖĶÉĶÚß@ĐŠDCsŌģĩŠĩŊß_ÁÜ°ÍŊYžÍÖģģîēÜļĖĩÄrégŖŦÖģĶĐĶĐĪŪĩÁŋĩÄTŧ°ûĸąģŧ¤ģî——ēÃ×ãŌÔĒĻÉŲÁŋĩÄÎĸÉúÎīČëĮÖÕߥŖÁíŌģˇŊÃæŖŦČįšû¸ĐČžĀÖØŖŦĸŽaÉúÔSļāđļˇŧ°ûŌō×ĶŖŦÔSļāäÍģ îŧ°ûĸąģŧ¤ģî˛ĸßwŌÆĩŊ¸ŊŊüĩÄÁÜ°ÍŊYŖŦÄŅĒŌēÖĐÕĐÄŧ¸üļāĩÄDCsŖŦŧ¤ģî¸üļāĩÄTŧ°ûĄŖŌō´ËŖŦäÍģ îŧ°ûļĖēÛÃüĩÄŌģŊYšûĘĮŖŦÔÚČÎēÎŊoļ¨rŋĖÁÜ°ÍŊYÖĐĩÄDCsĩÁŋĸˇ´ĶŗđļˇŦFöĩÄŽĮ°ĮérŖŦÃâŌߡ´ĒĩÄļČĸÅc¸ĐČžĩÄĀÖØŗĖļČŗÉąČĀũĄŖ
ËųŌÔŖŦäÍģ îŋšÔŗĘßfŧ°ûĘĮÉÚąøŧ°ûŖŦËüÔÚŊMŋÖвÉĶŋšÔĄŖČįšûĶĐČëĮÖŖŦDCsūąģŧ¤ģî˛ĸßwŌÆĩŊ¸ŊŊüĩÄÁÜ°ÍŊYĄŖÔÚÄĮĀīŖŦËüͨß^Īōδŧ¤ģîĩÄŨoÖúTŧ°ûēÍŧ°ûļžĐÔTÁÜ°Íŧ°ûŖ¨CTLsŖŠŗĘßfÔÚđļˇŦFöĘÕŧ¯ĩÄŋšÔíĸĶßmĒĐÔÃâŌߡ´ĒĄŖŧ¤ģîĩÄDCsÛÃüļĖēŖŦß@ĐŠŧ°ûĩÄŋėËŲ¸üĖæ´_ąŖÁËËü§ĩŊÁÜ°ÍŊYĩÄ“DÆŦ”˛ģā¸üĐÂĄŖ´ËÍâŖŦÄŊMŋÖĐÅÉĮ˛ĩÄäÍģ îŧ°ûĩÁŋēÍÕĐÄŧĩÄĖæ´úäÍģ îŧ°ûĩÄĩÁŋĸČĄQĶÚšĨôĩÄĀÖØŗĖļČĄŖŌō´ËŖŦÃâŌßĪĩŊyÄÜō¸ųūČëĮÖËųŗÉĩÄÎŖëUŗĖļČ×÷ŗöĪāĒĩġ´ĒĄŖÄãÄÜĪëĪķŗöŌģ¸üĮÉÃîĩÄĪĩŊyáŖŋÎŌ˛ģß@ôÕJéŖĄäÍģ îŧ°ûąģwîéĪČĖėÃâŌßĪĩŊyĩÄŌģ˛ŋˇÖŖŦŌōéËüĩÄĘÜķwĘĮ“Ķ˛ßBž”ĩÄŖŦ˛ģĪņBŧ°ûēÍTŧ°ûÄĮĶ“ŋÉßmĒ”ĄŖČģļøŖŦÕũČįÄãŦFÔÚŋĪļ¨ĀíŊâĩÄŖŦDCsëHÉĪŗäŽĪČĖėēÍßmĒĐÔĪĩŊyÖŽégĩÄ“ōÁē”ĄŖ
6.2.ŧ¤ģîĩÄžŪĘÉŧ°û
žŪĘÉŧ°ûŌ˛ĘĮÉÚąøŧ°ûŖŦËüĘØĐlÖøÎŌÉíķwąŠÂļĶÚÍâŊįĩÄ ^ĶōĄŖËüŋÉŌÔ×÷éĀŦģøĘÕŧ¯Æ÷ĄĸŋšÔŗĘßfŧ°ûģō´ÃÍĩÄĸĘÖ——ß@ČĄQĶÚËüËųĖÎĸhžŗĩÄĐÅĖĄŖÔÚėoÖš îBĪÂŖŦžŪĘÉŧ°ûÉÃéLÕûĀíŖŦĩĢÔÚŋšÔŗĘßfˇŊÃæ˛ĸ˛ģÉÃéLĄŖß@ĘĮŌōéžŪĘÉŧ°ûÖģĶĐÔÚąģIFN-γĩČđļˇŧ°ûŌō×Ķŧ¤ģîŖŦģōÕßËüĩÄÄŖĘŊ×ReĘÜķwąģČëĮÖ˛ĄÔķwŊYēĪēķŖŦ˛Åąíß_×ãōĩÄMHCēÍš˛´Ėŧ¤ˇÖ×ĶŌÔ×÷éŋšÔŗĘßfŧ°ûĄŖ
Ōō´ËŖŦžŪĘÉŧ°ûÅcäÍģ îŧ°ûĪāËÆŖŦËüÖģÔÚĶĐÎŖëUĩÄ|Î÷ĐčŌĒŗĘßfr˛ÅĶĐЧĩØŗĘßfŋšÔĄŖČģļøŖŦÖØŌĒĩÄĘĮŌĒÕJ×RĩŊäÍģ îŋšÔŗĘßfŧ°û˛ģĸČËŖŦžŪĘÉŧ°û˛ģÂÃĐĐĄŖëHÉĪŖŦDCsŋÉŌÔąģÃčĀLŗɲģ§ÎäÆ÷ĩÄ“zĶ°ĶÕß”ŖŦËûÅÄzđļˇĩÄŋėÕÕŖŦČģēķëxé_đöČĨĖáŊģËûĩÄķ¸æĄŖĪāąČÖŽĪÂŖŦžŪĘÉŧ°ûĘĮČĢ¸ąÎäŅbĩÄĘŋąøŖŦąØíÕžÁĸđļˇĄŖŽ žšŖŦžŪĘÉŧ°ûĘĮÎŌÔįÆÚˇĀĶųČëĮÖÕßĩÄÖ÷ŌĒÎäÆ÷ÖŽŌģĄŖąMšÜČį´ËŖŦËüČąˇĻÁ÷ĶĐÔĖáŗöÁËŌģĶĐȤĩÄî}ŖēČįšûŧ¤ģîĩÄžŪĘÉŧ°û˛ģÄÜÂÃĐĐĩŊĖÅŽTŧ°ûËųÔÚĩÄÁÜ°ÍŊYŖŦÄĮôŧ¤ģîĩÄžŪĘÉŧ°ûŗĘßfŋšÔĩÄÄÜÁĻĶĐĘ˛Ã´ĶÃÄØŖŋ´đ°¸ÔÚß@ĀīĄŖ
ŌģĩŠTŧ°ûąģäÍģ îŧ°ûŧ¤ģîŖŦËüžÍëxé_ÁÜ°ÍŊYŖŦͨß^ŅĒŌēŅhŖŦ˛ĸßMČë°lŅ×ĩÄŊMŋŌÔÍÖúđļˇĄŖČģļøŖŦß@ĐŠ“ĶĐŊōĩÄ”Tŧ°ûąØí˛ģāÖØĐ´Ėŧ¤ĄŖˇņtŖŦËüūÕJéđļˇŌŅŊŲĀûŖŦËüūģØĩŊėoÖš îBģōŌōēöŌļøËĀÍöĄŖß@žÍĘĮŧ¤ģîĩÄžŪĘÉŧ°ûĩÄĶÃÎäÖŽĩØĄŖÔÚŊMŋÖĐŖŦžŪĘÉŧ°ûŗ䎓ŧĶĶÍÕž”ŖŦąŖŗÖĶĐŊōĩÄTŧ°ûŧ¤ģîŖŦĘšËüŋÉŌÔĀ^Ām ĸÅcđļˇĄŖŌō´ËŖŦŗÉĘėĩÄäÍģ îŧ°ûŧ¤ģîδŧ¤ģîĩÄTŧ°ûŖŦļøŧ¤ģîĩÄŊMŋžŪĘÉŧ°ûÖ÷ŌĒšĻÄÜĘĮÖØĐ´Ėŧ¤ĶĐŊōĩÄTŧ°ûĄŖ
6.3.ŧ¤ģîĩÄBŧ°û
ĩÚČũˇNŋšÔŗĘßfŧ°ûĘĮŧ¤ģîĩÄBŧ°ûĄŖδŧ¤ģîĩÄBŧ°ûÔÚŋšÔŗĘßfˇŊÃæ˛ĸ˛ģÉÃéLŖŦŌōéËüÖģąíß_ĩÍËŽÆŊĩÄIIîMHCˇÖ×ĶēÍēÜÉŲģō]ĶĐB7ĄŖČģļøŖŦŌģĩŠBŧ°ûąģŧ¤ģîŖŦÆäąíÃæĩÄIIîMHCˇÖ×ĶēÍB7ĩ°°×ĩÄËŽÆŊžÍūōĄĐÔĩØÔöŧĶĄŖŌō´ËŖŦĶĐŊōĩÄBŧ°ûÄÜō×÷éThŧ°ûĩÄŋšÔŗĘßfŧ°ûĄŖBŧ°ûÔÚ¸ĐČžĩÄŗõĘŧëAļβģ×÷éAPCs°l]×÷ĶÃŖŦŌōéÄĮrËüßĘĮĖėÕæĩÄ——Ëüß]ĶĐąģŧ¤ģîĄŖČģļøŖŦÔÚ¸ĐČžß^ŗĖĩÄēķÆÚģōÔÚëSēķĩĸĐČžÖĐŖŦĶĐŊōĩÄBŧ°ûŗĘßfĩÄŋšÔ°l]ÖøÖØŌĒ×÷ĶÃĄŖß@ĘĮŌōéBŧ°ûąČÆäËûAPCsĶĐŌģžŪ´ķĩÄŨŖēBŧ°ûŋÉŌÔŧ¯ÖĐŋšÔŌÔšŠŗĘßfĄŖÔÚBŧ°ûĩÄĘÜķwÅcÆäĪāĒĩÄŋšÔŊYēĪēķŖŦBCRŧĶŋšÔĩÄÕûÍēĪÎīÄŧ°ûąíÃæąģŌÆŗũ˛ĸÍĪČëŧ°ûČĄŖÔÚÄĮĀīŖŦŋšÔąģĖĀíŖŦŅbŨdĩŊIIîMHCˇÖ×ĶÉĪŖŦ˛ĸąģß\ŨģØŧ°ûąíÃæŌÔšŠŗĘßfĄŖ
Bŧ°ûĘÜķwĻŋšÔžßĶиßĶHēÍÁĻŖŦŌō´ËËüĪņ“´ÅčF”ŌģĶŖŦĘÕŧ¯ŋšÔŌÔŗĘŦFŊoThŧ°ûĄŖĶÉĶÚÔÚThŧ°ûąģŧ¤ģîÖŽĮ°ąØíĶĐŌģļ¨ĩÁŋĩÄTŧ°ûĘÜķwąģŗĘŦFĩÄŋšÔŊģÂŖŦūšĀĶŖŦÔÚŋšÔĪāĻŨ^ÉŲĩÄĮérĪÂŖŦŧ¤ģîĩÄBŧ°ûąČÆäËûŋšÔŗĘßfŧ°ûŖ¨APCsŖŠÔÚŧ¤ģîŨoÖúTŧ°ûˇŊÃæžßĶĐ100ÖÁ10,000ąļĩÄŨĄŖBŧ°ûŗĘŦFŋšÔŌ˛ˇĮŗŖŅ¸ËŲĄŖÄBŧ°ûĘÜķw˛ļĢ@ŋšÔĩŊͨß^IIîMHCˇÖ×ĶÔÚŧ°ûąíÃæÕšĘžŋšÔŖŦ˛ģĩŊ°ëĐĄrĩÄrégžÍūß^ČĨĄŖŋÖŽŖŦŽĘ×´ÎĶöĩŊČëĮÖÕßrŖŦËųĶĐÄÜō×ReÔĖØļ¨ČëĮÖÕßĩÄBŧ°ûļŧĘĮδŊŧ¤ģîĩÄŖŦŌō´ËÖØŌĒĩÄAPCsĘĮŧ¤ģîĩÄäÍģ îŧ°ûĄŖČģēķŖŦÔÚđļˇßMĐĐĩÄÍŦrŖŦĮ°žĩÄŧ¤ģîžŪĘÉŧ°ûĪōđļˇÖĐĩÄTŧ°ûŗĘŦFŋšÔŖŦŌÔąŖŗÖËüĩÄģîÁĻĄŖÔÚ¸ĐČžēķÆÚŖŦģōÕßČįšûÔŲ´ÎĶöĩŊĪāÍŦĩÄČëĮÖÕßŖŦŊōØS¸ģĩÄBŧ°ûĘĮˇĮŗŖÖØŌĒĩÄAPCs——ŌōéËüŋÉŌÔͨß^ŧ¯ÖĐÉŲÁŋŋšÔßMĐĐŗĘŦFŖŦŅ¸ËŲŧ¤ģîŨoÖúTŧ°ûĄŖ
7.IîMHCŗĘßfĩÄßŨ
ŌĒÕæÕũĀíŊâéĘ˛Ã´ŋšÔŗĘßfĘĮ´ķ×ÔČģ×îĨ´ķĩÄ“°lÃ÷”ÖŽŌģŖŦÎŌĐčŌĒÉÔÎĸËŧŋŧŌģĪÂß@Ōģķ@ČËģîĶąŗēķĩÄßŨĄŖĘ×ĪČŖŦÎŌĐčŌĒŌģî}ŖēéĘ˛Ã´ŌĒßMĐĐMHCŗĘßfŖŋéĘ˛Ã´˛ģ×Tŧ°ûĘÜķwĪņBŧ°ûĘÜķwÄĮĶ×ReδŗĘßfĩÄŋšÔŖŋß@ĘĮŌģÕæÕũĩÄɡÖî}ŖŦŌōéÎŌĶÕĩÄĘĮɡNˇĮŗŖ˛ģÍŦĩÄÕšĘžŖēIîēÍIIîĄŖËųŌÔ×ÎŌÖđŌģz˛éß@ĐŠî}ĄŖ
IîŗĘßfĩÄŌģÔŌōĘĮĸĸĘÖTŧ°ûĩÄ×ĸŌâÁĻŧ¯ÖĐÔÚ¸ĐČžĩÄŧ°ûÉĪŖŦļø˛ģĘĮŅĒŌēÖĐēÍŊMŋÖĐĩIJĄļžēÍÆäËû˛ĄÔķwÉĪĄŖÖģŌĒ˛ĄÔķwÍŖÁôÔÚÎŌĩÄŧ°ûÍâŖŦŋšķwžÍŋÉŌÔËĶËüŌÔšŠŖIÍĖĘÉŧ°ûÆÆÄŖŦ˛ĸŋÉŌÔÅcËüŊYēĪ˛ĸ×čÖšËüŌũ°l¸ĐČžĄŖĶÉĶÚÃŋ{ŧ°ûBŧ°ûÃŋÃëŋÉŌÔąÃŗöŧs2,000ŋšķwˇÖ×ĶŖŦß@ĐŠŋšķwĘĮ“ąãŌË”ĩÄÎäÆ÷ŖŦĻŧ°ûÍâČëĮÖÕߡĮŗŖĶĐЧĄŖČģļøŖŦŌģĩŠÎĸÉúÎīßMČëŧ°ûŖŦŋšķwžÍoˇ¨Ķ|ŧ°ËüĄŖŽß@ˇNĮér°lÉúrŖŦžÍĐčŌĒĸĘÖTŧ°û——ŖéTÔOĶĶÃíÆÆĸĐČžŧ°ûĩĸßŋÆŧŧĄĸ“°ēŲF”ĩÄÎäÆ÷ĄŖĸĘÖTŧ°ûĐčŌĒ×Re¸ĐČžŧ°ûÉĪĶÉIîMHCˇÖ×ĶŗĘßfĩÄŋšÔŖŦß@´_ąŖÁËCTL˛ģūĀËŲMrégČĨšĨôŧ°ûÍâĩÄČëĮÖÕß——ß@ĐŠČëĮÖÕßͨŗŖŋÉŌÔąģŋšķwˇĮŗŖĶĐЧĩØĖĀíĄŖ
´ËÍâŖŦ×δŗĘßfĩÄŋšÔĐÅĖTŧ°ûĸûĸOÆäÎŖëUĄŖĪëĪķŌģĪÂŖŦČįšûδ¸ĐČžĩÄŧ°ûÅöĮÉĶĐËĀ˛ĄļžĩÄËéÆŦÕŗ¸ŊÔÚËüĩÄąíÃæŖŦĸĘÖTŧ°û×ReÁËß@ˇNδŗĘßfĩÄŋšÔ˛ĸĸËĀÁËÄĮĐŠošŧĩÄÅÔĶ^Õßŧ°ûĄŖÄĮŽČģ˛ģĐĐĄŖ
IîÕšĘžÖŽËųŌÔČį´ËÖØŌĒŖŦÁíŌģÔŌōĘĮ´ķļāĩÔÚ˛ĄÔķw¸ĐČžĩÄŧ°ûÖĐŽaÉúĩÄĩ°°×Ų|ļŧÁôÔÚŧ°ûČŖŦÄδĩŊß_ŧ°ûąíÃæĄŖŌō´ËŖŦ]ĶĐIîÕšĘžŖŦÔSļā˛ĄÔķw¸ĐČžĩÄŧ°ûĸ˛ģūąģĸĘÖTŧ°ûzyĩŊĄŖĘÂÉĪŖŦIîMHCÕšĘžĩÄÉņÆæÖŽĖÔÚĶÚŖŦÔtÉĪŖŦÃŋČëĮÖ˛ĄÔķwĩÄĩ°°×Ų|ļŧŋÉŌÔąģĮĐËé˛ĸĶÉIîMHCˇÖ×ĶÕšĘžŊoĸĘÖTŧ°û˛éŋ´ĄŖ
×îēķŖŦĶÉĶÚËüĩÄĘÜķw×ReδąģÆŦļÎģ¯ēÍŗĘßfĩÄ“ĖėČģ”ŋšÔŖŦBŧ°ûëHÉĪĖĶÚÁĶŨĄŖÔŌōĘĮ´ķļāĩĩ°°×Ų|ąØíÕÛ¯B˛ÅÄÜÕũŗŖ°l]×÷ĶÃĄŖĶÉĶÚß@ˇNÕÛ¯BŖŦÔSļāBŧ°ûĘÜķwŋÉÄÜ×ReĩÄąíÎģoˇ¨˛éŋ´——ŌōéËüÎģĶÚÕÛ¯Bĩ°°×Ų|ˇÖ×ĶĩÄȲŋĄŖĪāąČÖŽĪÂŖŦŽĩ°°×Ų|ąģĮĐËéŗÉļĖÆŦļβĸĶÉIîMHCˇÖ×ĶŗĘßfrŖŦąíÎģoˇ¨ĻĸĘÖTŧ°ûë[˛ØĄŖ
Ōō´ËŖŦIîMHCŗĘßfĩÄßŨēÜČŨŌ×ĀíŊâŖŦĩĢéĘ˛Ã´MHCˇÖ×ĶČį´ËļāBĐÔŖŋŽ žšŖŦÔÚČËîČēķwÖĐ´æÔÚČį´ËļāĩIJģÍŦĐÎĘŊŖŦŌÔÖÁĶÚÎŌ´ķļāĩČËĀ^ŗĐÁËÁųˇN˛ģÍŦĩÄIîˇÖ×ĶĩÄģųŌōĄŖß@ŋ´ÆđíĶĐücß^ˇÖŖŦ˛ģĘĮáŖŋ
ēðÉŖŦŧŲÔOÖģĶĐÉŲĩסN˛ģÍŦĩÄIîMHCĩ°°×ĄŖŦFÔÚĪëĪķŌģĪÂŖŦČįšû˛Ąļž°lÉúÍģ×ŖŦŌÔÖÁĶÚËüĩÄëÄļÎoˇ¨Åcß@ĐŠMHC IˇÖ×ĶÖĐĩÄČÎēÎŌģŊYēĪĄŖß@ĶĩIJĄļžŋÉÄÜūĪûįÕûČËîČēķwŖŦŌōé]ĶĐĸĘÖTŧ°ûŋÉŌÔąģŧ¤ģîíÆÆIJĄļž¸ĐČžĩÄŧ°ûĄŖŌō´ËŖŦļāBĐÔMHCˇÖ×ĶÖÁÉŲŊoČēķwÖĐĩÄŌģĐŠČËĖᚊÁËÔÚŊÆģĢ˛ĄÔķwšĨôÖĐÉú´æĩÄCūĄŖ
ēÃĩÄŖŦĩĢÎŌéĘ˛Ã´ūĶĐÁųˇNIîMHCĩ°°×ĩÄģųŌōÄØŖŋß@ËÆēõēÜļāŖŦĖØeĘĮŋŧ]ĩŊIîMHCĩ°°×ĘĮČį´ËĩÄļāBĐÔĄŖ´đ°¸ĘĮ“íĶĐ”ļāß_ÁųˇN˛ģÍŦĩÄIîMHCˇÖ×ĶĩÄŋÉÄÜĐÔÔöŧĶÁËÎŌÃŋČËŖŦķwļøŅÔŖŦÖÁÉŲĶĐŌģˇNIîMHCˇÖ×ĶŋÉŌÔßmĒŊoļ¨˛ĄÔķwĩÄĩ°°×Ų|ÆŦļÎĩĸÅÂĘĄŖĘÂÉĪŖŦ¸ĐČžHIV-1˛ĸĀ^ŗĐÁË×î´ķĩÁŋ˛ģÍŦIîMHCˇÖ×ĶŖ¨ÁųˇNŖŠĩÄČËÆŊžųÛÃüÃ÷ī@éLĶÚÖģĶĐÎåˇNģō¸üÉŲ˛ģÍŦIîˇÖ×ĶģųŌōĩÄķwĄŖß@ĀīĩÄËŧŋŧĘĮŖŦëSÖøHIV-1ĩÄÍģ×ŖŦíĶиüļāĩIJģÍŦIîˇÖ×ĶÔöŧĶÁËÍģײĄļžĩ°°×ŋÉŌÔąģŗĘßfĩĸÅÂĘĄŖéĘ˛Ã´ĘĮÁųˇNŖŦļø˛ģĘĮĘŽˇNIîMHCˇÖ×ĶĩÄģųŌōŖŋÎŌ˛ģÖĒĩĀŖĄ
8.IIîMHCŗĘßfĩÄßŨ
ēÃĩÄŖŦIîMHCŗĘßfˇĮŗŖĶĐŌâÁxĄŖĩĢĘĮIIîŗĘßfÄØŖŋÕ§Ōģŋ´ŖŦß@ˇNëpÖØÕšĘžŖ¨IîēÍIIîŖŠŋÉÄÜŋ´Æđíß^ĶÚÍësĄŖČģļøŖŦąØíĀíŊâĩÄĘĮŖŦÔSļā˛ĄÔķw˛ģ¸ĐČžČËîŧ°ûŖēËüēܸßÅdÔÚÎŌĩÄŧ°ûÍâÔÚÎŌĩÄŊMŋģōŅĒŌēÖĐÉúģîē͡ąÖŗĄŖČįšûŋšÔŗĘßfŧ°ûÖģÄÜÕšĘžËü¸ĐČžĩIJĄÔķwŽaÉúĩÄĩ°°×Ų|ŖŦÄĮôęPĶÚÔSļā×îÎŖëUÎĸÉúÎīĩÄĮéķžÍĶĀßh˛ģūĩŊß_ÁÜ°ÍŊYÖĐĩÄÖ¸]ÖĐĐÄĄŖͨß^ĘšĶÃIIîMHCˇÖ×ĶÔÚĮ°žĐû÷ŋhžŗĩÄĶąžŖŦŋÉŌÔĪōŨoÖúTŧ°ûĖᚊęPĶÚËųĶĐîĐÍČëĮÖÕßĩÄĮéķĄŖ
ĩĢĘĮŨoÖúTŧ°û˛ģÄÜÖģĘĮ×ReδŗĘßfĩÄŋšÔáŖŋŽ žšŖŦËü˛ģĘĮĸĘÖŖŦËųŌÔ]ĶĐÅÔĶ^ÕßĸžĩÄî}ĄŖŽČģŖŦß@ĘĮÕæĩÄŖŦĩĢß@ĀīČÔČģĶĐŌģ°˛ČĢî}ĄŖŋšÔŗĘßfŧ°ûÖģÔÚđļˇßMĐĐr˛ÅĶĐЧĩØŗĘßfŋšÔŖŦŨoÖúTŧ°ûąģēYßxŌÔ´_ąŖËü˛ģūĻÎŌ×ÔŧēĩÄĩ°°×Ų|ŽaÉúˇ´ĒĄŖŌō´ËŖŦŨoÖúTŧ°ûēÍŋšÔŗĘßfŧ°ûļŧąØí“ÍŦŌâ”°lÉúÁËČëĮÖŖŦŨoÖúTŧ°û˛ÅÄÜąģŧ¤ģîĄŖËųŌÔŖŦŨoÖúTŧ°ûÖģ×ReŗĘßfĩÄŋšÔĩÄŌĒĮķ´_ąŖÁ˲ŋĘđÔÚÖÂÃüĩÄßmĒĐÔÃâŌßĪĩŊyĩÄQļ¨˛ģĘĮĶÉÎŧ°û×öŗöĩÄĄŖ
ÍŦĶŖŦĪņIîˇÖ×ĶŌģĶŖŦIIîˇÖ×ĶŗĘßfĩ°°×Ų|ĩÄĐĄÆŦļÎĄŖŌō´ËŖŦÔÚŗĘßfÆÚégŨoÖúTŧ°ûŋÉŌÔ“ŋ´ĩŊ”ĩÄÄŋËĩÁŋßhßhŗŦß^ÔÚ´ķĐÍÕÛ¯Bĩ°°×Ų|ÖĐŋÉšŠ˛éŋ´ĩÄĩÁŋĄŖß@ˇNU´ķĩÄÄŋËĩÁŋĩÄēķšûĘĮ¸üÁŌĄĸ¸üļāĶģ¯ĩÄÃâŌߡ´ĒŖŦÔSļā˛ģÍŦĩÄŨoÖúTŧ°ûĸąģŧ¤ģî——ŨoÖúTŧ°ûĩÄĘÜķw×ReŗÉČëĮÖÕßŋšÔĩÄÔSļā˛ģÍŦĩÄąíÎģĄŖ
9.Ŋģ˛æŗĘßf
ąMšÜIîēÍIIîÍžŊĩġÖëxĘĮ“Ōt”ŖŦĩĢŌŅŊī@ĘžŖŦÄŗĐŠČēĩÄŋšÔŗĘßfŧ°ûÄÜōzČĄÍâÔ´ĐÔŋšÔ˛ĸĸËüËÍČëIîÍžŊŖŦŌÔąãĶÉIîMHCˇÖ×ĶŗĘßfĄŖß@ˇNß`ˇ´IîÕšĘžĩġĮˇ¨ĘšĶÃąģˇQéŊģ˛æŗĘßfĄŖÆäĀíÄîĘĮŖŦČįšûŌģÂÃ÷ĩIJĄÔķwŖ¨ĀũČįŖŦ˛ĄļžŖŠÕŌĩŊÁËąÜÃâ¸ĐČžŋšÔŗĘßfŧ°ûĩġŊˇ¨ŖŦĩĢČÔČģÄÜÔÚÉíķwĩÄÆäËûŧ°ûÖиĐČžē͡ąÖŗŖŦŊģ˛æŗĘßfĸŊoÃâŌßĪĩŊyŌģŧ¤ģîáĻÔ˛ĄÔķwĩÄCTLsĩÄCūĄŖĩŊÄŋĮ°éÖšŖŦŋØÖÆŊģ˛æŗĘßfĩÄŌtēÍÉæŧ°ĩÄCÖÆÉĐδÃ÷´_ļ¨ÁxĄŖ´ËÍâŖŦÉвģĮåŗūÄAPCÍâ˛ŋzČĄĩÄŋšÔĶÉIîMHCˇÖ×ĶŊģ˛æŗĘßfĻÕũŗŖČËķwÃâŌßĪĩŊyĩÄšĻÄÜĘĮˇņÖØŌĒĄŖ
10.ˇĮŊĩäMHCˇÖ×ĶēÍÖŦŲ|ŗĘßf
IîēÍIIîMHCˇÖ×ĶąģˇQéŊĩäMHCˇÖ×ĶĄŖŌō´ËŖŦÕũČįÄãËųÄÜîAÁĪĩÄŖŦŌ˛´æÔÚˇĮŊĩäMHCˇÖ×ĶĄŖÆäÖĐŅĐžŋĩÃ×îēÃĩÄĘĮCD1ĩ°°×ŧŌ×åĄŖß@ĐŠˇĮŊĩäMHCˇÖ×ĶÔÚŊYÉĪîËÆĶÚIîMHCˇÖ×ĶŖŦËüĶÉŌģéLĩÄÖØæĩ°°×ŊMŗÉŖŦÅcβ2-ÎĸĮōĩ°°×ÅäĻĄŖČģļøŖŦÅcžßĶĐßmēĪŊYēĪļĖëÄĩÄ°ŧ˛ÛĩÄŊĩäMHCˇÖ×Ķ˛ģÍŦŖŦCD1ˇĮŊĩäMHCˇÖ×ĶžßĶĐŊYēĪÖŦŲ|ĩÄÕũ´_ĐÎ îĄŖCD1ˇÖ×ĶŋÉŌÔÄŧ°ûČĩIJģÍŦ ^Ķō“˛ÉĶ”ÖŦŲ|ŖŦ˛ĸÄÜÔÚŋšÔŗĘßfŧ°ûĩÄąíÃæŗĘßfß@ĐŠˇÖ×ĶŖŦÄļøŧ¤ģîTŧ°ûĄŖŌō´ËŖŦČËÍÆyß@ĐŠˇĮŊĩäMHCˇÖ×ĶéTŧ°ûĖᚊÁËŌģˇNz˛éŧ°ûÖŦŲ|ŊMŗÉĩÄÍžŊŖŦžÍĪņIîMHCˇÖ×ĶÔĘÔSTŧ°ûz˛éŧ°ûĩÄĩ°°×Ų|ŌģĶĄŖ
ÔÚÃâŌßWÖĐŖŦËÆēõÃŋlŌtļŧĶĐŌģĀũÍâŖŦß@ĀīĩÄŌtĘĮTŧ°ûÖģ×ReĶÉIîēÍIIîMHCˇÖ×ĶŗĘßfĩÄĩ°°×Ų|ÆŦļÎĄŖī@ČģŖŦCD1ĪōTŧ°ûŗĘßfÖŦŲ|ĘĮĻß@ŌģŌtĩÄĀũÍâĄŖČģļøŖŦÄŋĮ°ÉвģĮåŗūÖŦŲ|ŗĘßfĻÃâŌߡĀĶųĩÄÖØŌĒĐÔĄŖŌō´ËŖŦÎŌĸ“ÔŗÖŌt”ŖŦŧ´Tŧ°ûÖģ×Reĩ°°×Ų|ŋšÔĄŖČģļøŖŦÕ×ĸŌâŖŦëSÖøĻCD1ŗĘßfĩÄÖŦŲ|ßMĐиüļāŅĐžŋŖŦß@ˇNĮérŋÉÄÜū¸ÄץŖ
11.MHCĩ°°×ēÍÆ÷šŲŌÆÖ˛
ŗũÁËÔÚŋšÔŗĘßfÖĐĩÄ×ÔČģ×÷ĶÃÍâŖŦMHCˇÖ×ĶÔÚÆ÷šŲēÍŊMŋŌÆÖ˛ĩġĮ×ÔČģhžŗÖĐŌ˛ēÜÖØŌĒĄŖŌÆÖ˛ŅĐžŋëHÉĪĘŧĶÚ20ĘĀŧo30Äę´úŖŦÉæŧ°ĐĄĘķÄ[ÁöĩÄōĄŖÔÚÄĮĐŠČÕ×ĶĀīŖŦÄ[ÁöͨŗŖĘĮͨß^ÔÚĐĄĘķƤÄwÉĪÄĻ˛ÁŌģĐŠŋÉÅÂĩÄģ¯WÎīŲ|ŖŦČģēķĩČ´ũēÜéLrég˛ÅÄÜ°lÕšŗöÄ[ÁöĄŖĶÉĶÚÖÆÔėß@ĐŠÄ[ÁöˇĮŗŖÂéŠŖŦÉúÎīWŧŌĪŖÍûÔÚĐĄĘķËĀÍöēķÄÜąŖŗÖÄ[Áöŧ°û´æģîŌÔšŠŅĐžŋĄŖËûͨß^ĸŌģĐŠÄ[Áöŧ°û×ĸÉäĩŊÁíŌģÖģŊĄŋĩĩÄĐĄĘķķwČíŦFß@ŌģücŖŦÔÚÄĮĀīŧ°ûĸĀ^ĀmÉúéLĄŖČģļøŖŦËûĶ^˛ėĩŊŖŦÖģĶĐÔÚšŠķwēÍĘÜķwí×ÔÍŦŌģŊüŊģĪĩĐĄĘķrŖŦÄ[Áöŧ°û˛ÅÄÜŗÉšĻŌÆÖ˛ĄŖļøĮŌŖŦÆˇĪĩÔŊŊüŊģŖŦŌÆÖ˛´æģîĩÄCūÔŊēÃĄŖß@éÔėÔSļāŊņĖėÃâŌßWŧŌŌĀŲĩÄŊüŊģĐĄĘķÆˇĪĩĖᚊÁËĶÁĻĄŖ
ŌģĩŠŊüŊģĐĄĘķÆˇĪĩŋÉĶÃŖŦÃâŌßWŧŌé_ĘŧŅĐžŋĸÕũŗŖŊMŋÄŌģĘķËŌÆÖ˛ĩŊÁíŌģĘķËĄŖËûÁĸŧ´×ĸŌâĩŊŖŦČįšûĸŌģĐĄKƤÄwÄŌģÖģĐĄĘķŌÆÖ˛ĩŊÁíŌģÖģĐĄĘķĩÄƤÄwÉĪŖŦÖģŌĒÉÖģĐĄĘķí×ÔÍŦŌģŊüŊģÆˇĪĩŖŦĐÂƤÄwžÍąŖŗÖŊĄŋĩĩġÛŧtÉĢ˛ĸĀ^ĀmÉúéLĄŖĪāąČÖŽĪÂŖŦŽß@ōÔÚˇĮŊüŊģĐĄĘķÉĪßMĐĐrŖŦŌÆÖ˛ĩÄƤÄwÔÚ×ĐĄrČ×°×Ŗ¨ŌōéŅĒŌēšŠĒąģĮĐāŖŠ˛ĸĮŌŋĘĮËĀÍöĄŖÃâŌßWŧŌÕJéß@ˇNÁĸŧ´ĩÄŌÆÖ˛ÅÅŗâŌģļ¨ĘĮĶÉĶÚÄŗˇNßz÷˛ģŧæČŨĐÔŖŦŌōéËü˛ģū°lÉúÔÚžßĶĐĪāÍŦģųŌōĩÄŊüŊģĐĄĘķÉíÉĪĄŖéÁË´_ļ¨Éæŧ°ŊMŋŧæČŨĐÔŖ¨ŊMŋĪāČŨĐÔŖŠĩÄģųŌōŖŦÃâŌßWŧŌÅāĶũĐĄĘķŖŦÔėÁËÖģĶĐ×ģųŌō˛ģÍŦĩÄÆˇĪĩŖŦĩĢß@ĐŠÆˇĪĩČÔČģ˛ģßmĶÃĶÚŊMŋŌÆÖ˛ĄŖÃŋŽËûßMĐĐß@ĐŠōrŖŦËû˛ģā×ReŗöŌģŊMÎģĶÚĐĄĘķ17ĖČžÉĢķwÉĪĩÄÍësģųŌō——Ëû×îŊKˇQÖŽé“Ö÷ŌĒŊMŋĪāČŨĐÔÍēĪķw”ģōMHCĄŖ
Ōō´ËŖŦÎŌÔÚŋšÔŗĘßfąŗž°ĪÂĶÕĩÄMHCˇÖ×ĶÕũĘĮ§ÖÂŌÆÖ˛Æ÷šŲÁĸŧ´ÅÅŗâĩġÖ×ĶĄŖŊYšû°lŦFŖŦĸĘÖTŧ°ûĻ“Íâí”ĩÄMHCˇÖ×ĶĖØeÃô¸ĐŖŦŽËüŋ´ĩŊß@ĐŠˇÖ×ĶrŖŦËüūšĨô˛ĸĸËĀąíß_ß@ĐŠˇÖ×ĶĩÄŧ°ûĄŖËü×îĪ˛gĩÄÄŋËÖŽŌģĘĮŗÉžčŲÆ÷šŲČŅĒšÜĩÄŧ°ûĄŖͨß^ÆÆÄß@ĐŠŅĒšÜŖŦCTLsĮĐāÁËŌÆÖ˛Æ÷šŲĩÄŅĒŌēšŠĒŖŦͨŗŖ§ÖÂÆäËĀÍöĄŖŌō´ËŖŦŌÆÖ˛ÍâŋÆátÉúÔDÆĨÅäžßĶĐĪāÍŦMHCˇÖ×ĶĩÄšŠķwēÍĘÜķwĄŖČģļøŖŦÕŌĩŊß@ĶĩÄÆĨÅäēÜĀ§ëyĄŖĘÂÉĪŖŦūšĀĶŖŦČįšûÄãÄÜōŊĶĶ|ĩŊ1000ČfÅcÄãoęPĩÄķwžčĢIĩÄÆ÷šŲŖŦÄãÕŌĩŊÅcÄãËųĶĐIîēÍIIîMHCˇÖ×ĶÍęČĢÆĨÅäĩÄCūÖģĶĐ´ķŧs50%ĄŖŌō´ËŖŦMHCˇÖ×ĶĩÄļāĶĐÔŖŦÔÚąŖ×oÎŌÃâĘÜĐÂĩÄģōÍģ×ČëĮÖÕßĮÖēĻˇŊÃæˇĮŗŖÖØŌĒŖŦéÆ÷šŲŌÆÖ˛ÔėÁËŌģÕæÕũĩÄî}ĄŖī@ČģŖŦÃâŌßĪĩŊy˛ĸ]ĶĐŋŧ]ĩŊÆ÷šŲŊģQĩÄŅŨ×ŖĄ
12.ŋŊYD
Äúū×ĸŌâĩŊŖŦÎŌĩÄÕĒŌĒDŦFÔÚ°üĀ¨ŋšÔŗĘßfŧ°ûŧ°ÆäMHCēÍB7ˇÖ×ĶĄŖ
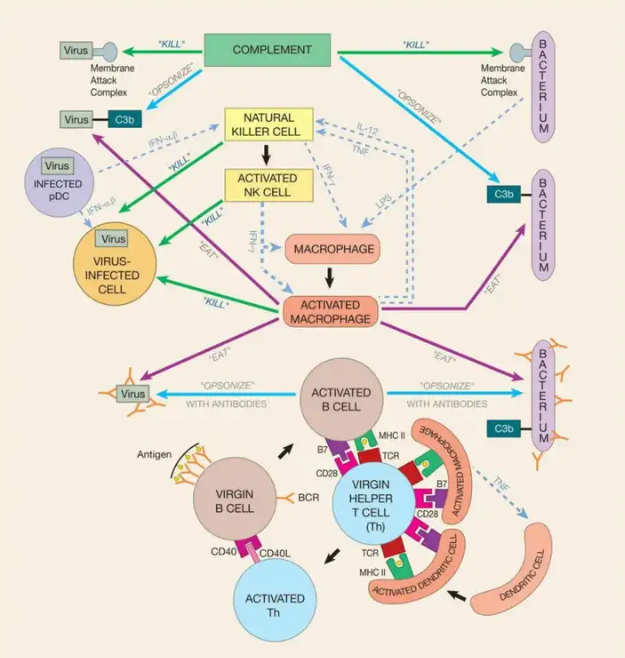
ģØî
IîMHCˇÖ×ĶŗäŽÕšĘžÅÆŖŦÕšĘžŧ°ûȲŋ°lÉúĩÄĮérĄŖĀũČįŖŦŽ˛ĄļžĮÖČžŧ°ûrŖŦËüūĀûĶÃÔŧ°ûĩÄÉúÎīēĪŗÉCÖÆíŽaÉú˛Ąļžĩ°°×ĄŖß@ĐŠĩ°°×Ų|ÖĐĩÄŌģĐŠąģĩ°°×øķwĮиîŗÉĐĄÆŦļÎŖ¨ëÄŖŠŖŦ˛ĸĶÉTAPŪDß\ĩ°°×y§ßMČëČŲ|žWŖ¨ERŖŠĄŖÔÚÄĮĀīŖŦëÄąģIîˇÖ×Ķ“ÃæÔ”ĄŖÄĮĐŠéLļČ´ķŧséžÅ°ąģųËáŖŦ˛ĸÔÚÆäÄŠļËžßĶĐßmŽ°ąģųËáĩÄëÄūąģŊYēĪÔÚIîMHCˇÖ×ĶĩÄ°ŧ˛ÛÖĐŖŦ˛ĸąģß\ŨĩŊŧ°ûąíÃæĄŖͨß^ßÃčÄĮĀīÕšĘžĩÄMHC I-ëÄÍēĪÎīŖŦĸûĐÔTŧ°ûŋÉŌÔ¸QŌŧ°ûȲŋŖŦŌÔ´_ļ¨ËüĘĮˇņąģ¸ĐČž˛ĸĒÔąģ´Ũ§ĄŖIIîMHCˇÖ×ĶŌ˛ĘĮV¸æÅÆŖŦĩĢËüąģÔOĶĶÃíĖáĐŅŨoÖúTŧ°ûÕũÔÚßMĐĐđļˇĄŖIIîˇÖ×ĶÔÚERÖĐŊMŅbŖŦžÍĪņIîˇÖ×ĶŌģĶŖŦĩĢĶÉĶÚ˛ģ×æĩ°°×ÕŧūÁËËüĩÄŊYēĪ°ŧ˛ÛŖŦIIîˇÖ×Ķ˛ģÔÚERÖĐėĘ°ëÄĄŖĪāˇ´ŖŦIIî-˛ģ×æÍēĪÎīąģß\ŨŗöER˛ĸßMČëÁíŌģˇQéČķwĩÄŧ°û ^ĶōĄŖÔÚÄĮĀīŖŦËüÅcͨß^ÍĖĘÉ×÷ĶÃßMČëŧ°ûĩÄĩ°°×Ų|ĪāĶöŖŦ˛ĸąģøĮиîŗÉëÄĄŖß@ĐŠëÄëSēķČĄ´úÁËŌģÖąĘØ×oIIîˇÖ×Ķ°ŧ˛ÛĩIJģ×æŖŦMHC-ëÄÍēĪÎīąģß\ŨĩŊŧ°ûąíÃæŖŦšŠThŧ°ûÕšĘžĄŖͨß^ß@ˇNĮÉÃîĩÄCÖÆŖŦIIîˇÖ×ĶėĘ°í×Ôŧ°ûÍâĩÄĩ°°×Ų|ŅÜÉúĩÄëÄŖŦĩĢąÜÃâÁËí×Ôŧ°ûČÖÆÔėĩÄĩ°°×Ų|ĩÄëÄĄŖMHCˇÖ×ĶÕšĘžĩ°°×Ų|ÆŦļÎąČÕšĘžÍęÕûĩ°°×Ų|ĶĐ×ŨĄŖĘ×ĪČŖŦ´ķļāĩ˛Ąļžĩ°°×ͨŗŖë[˛ØÔÚąģ¸ĐČžĩÄŧ°ûȲŋŖŦ˛ģūŗöŦFÔÚŧ°ûąíÃæĄŖŌō´ËŖŦŗũˇĮąģIîMHCˇÖ×ĶĐû÷ŖŦˇņtß@ĐŠĩ°°×Ų|ĶĀßh˛ģūąģĸûĐÔTŧ°ûŋ´ĩŊĄŖ´ËÍâŖŦĶÉĶÚĩ°°×Ų|ÕÛ¯BŋÉŌÔë[˛Øĩ°°×Ų|ĩÄ´ķ˛ŋˇÖŖŦĸĩ°°×Ų|ĮиîŗÉĐĄëÄŋÉŌÔŊŌĘžÔSļāÔÚÍęÕûĩ°°×Ų|ÖĐoˇ¨ŊĶĶ|ĩÄÔÚTŧ°û°ĐËĄŖŌō´ËŖŦMHCÕšĘž´ķ´ķÔöŧĶÁËCTL×Reąģ¸ĐČžŧ°ûēÍŨoÖúTŧ°ûąģĖáĐŅÎĸÉúÎīšĨôĩÄŋÉÄÜĐÔĄŖIîēÍIIîMHCˇÖ×ĶļŧˇĮŗŖļāBĐÔŖŦČËîíĶĐß@ÉîMHCˇÖ×ĶĩÄļāģųŌōĄŖŌō´ËŖŦÄãĩÄMHCˇÖ×ĶēÜŋÉÄÜÄÜōÕšĘží×Ô´ķļāĩ˛ĄÔķwĩÄëÄŖŦ˛ĸĮŌÖÁÉŲĶĐŌģĐŠČËČēÖĐĩÄČËíĶĐÄÜōÕšĘží×ÔČÎēβĄÔķwĩÄëÄĩÄMHCˇÖ×ĶĄŖŋšÔŗĘßfŧ°ûĘĮĖØĘâĩÄÃâŌßĪĩŊyŧ°ûŖŦŋÉŌÔĖᚊIîēÍIIîMHCÕšĘžŌÔŧ°š˛´Ėŧ¤ĄŖÔÚšĨôĩÄŗõĘŧëAļÎ×îÖØŌĒĩÄŋšÔŗĘßfŧ°ûĘĮäÍģ îŧ°ûŖŦŌōéß@ˇNŧ°ûŋÉŌÔŧ¤ģîÔĘŧTŧ°ûĄŖŽDCÔÚđöŦFözyĩŊÎŖëUĐÅĖrŖŦËüé_ĘŧŗÉĘėŖŦ˛ĸy§ÆäđļˇŋšÔĩÄØÎīßwŌÆĩŊ¸ŊŊüĩÄÁÜ°ÍŊYĄŖÔÚÄĮĀīŖŦäÍģ îŧ°ûĘšĶÃIIîMHCˇÖ×ĶÕšĘžËüÔÚŊMŋÖĐĘÕŧ¯ĩÄĩ°°×Ų|ÆŦļÎŖŦĘšĶÃIîMHCˇÖ×ĶÕšĘžŋÉÄÜÔÚđöŦFö¸ĐČžäÍģ îŧ°ûĩIJĄļžģōŧžúÖÆÔėĩÄĩ°°×Ų|ÆŦļÎĄŖͨß^ß@ˇNˇŊĘŊŖŦäÍģ îŧ°ûĶĐЧĩØÅÄzÁËĮ°žÕũÔÚ°lÉúĩÄĘÂĮéĩÄŋėÕÕŖŦĸÆä§ĩŊTŧ°ûØS¸ģĩÄĩØˇŊŖŦČģēķßMĐĐ“ÕšĘžēÍÖvĘö”ŖŦŌÔŧ¤ģîTŧ°ûĄŖ
žŪĘÉŧ°ûÔÚąģÎŖëUĐÅĖŧ¤ģîēķŖŦŌ˛ŋÉŌÔ×÷éŋšÔŗĘßfŧ°ûĄŖČģļøŖŦŧ¤ģîĩÄžŪĘÉŧ°û˛ģūĮ°ÍųÁÜ°ÍŊYŗĘßfŋšÔĄŖËüÁôÔÚŊMŋÖĐÅcČëĮÖÕßđļˇĄŖŌō´ËŖŦžŪĘÉŧ°ûÔÚßmĒĐÔÃâŌßĪĩŊyąģŧ¤ģîēķŗĘßfŋšÔ×îĶĐĶÃĄŖÄĮrŖŦŊMŋÖĐŧ¤ģîĩÄžŪĘÉŧ°ûŋÉŌÔąŖŗÖŊōØS¸ģĩÄTŧ°ûĩÄģîĐÔŖŦŅĶéLËüĶĐЧĻŋšČëĮÖÕßĩÄrégĄŖŧ¤ģîĩÄBŧ°ûĘĮĩÚČũˇNŋšÔŗĘßfŧ°ûŖŦĩĢÍŦĶŖŦß@ĐŠŧ°ûÔÚĸĶĻĐÂČëĮÖÕßĩÄßmĒĐÔˇ´ĒÖвĸ˛ģĶĐĶÃĄŖÔŌōĘĮBŧ°ûÔÚÄÜ×÷éŋšÔŗĘßfŧ°ûÖŽĮ°ŖŦąØíĪČąģŨoÖúTŧ°ûŧ¤ģî——ļøThŧ°ûąØíĩČ´ũąģäÍģ îŧ°ûŧ¤ģîĄŖŌō´ËŖŦBŧ°ûÖąĩŊßmĒĐÔÃâŌߡ´ĒŌŅŊĸĶēķ˛Å“Ģ@ĩÃÕJ×C”ŗÉéŋšÔŗĘßfŧ°ûĄŖąMšÜČį´ËŖŦŌģĩŠąģŧ¤ģîŖŦBŧ°ûąČDCēÍžŪĘÉŧ°ûĶĐŌģžŪ´ķĩÄŨŖēBŧ°ûŋÉŌÔĘšĶÃËüĩÄĘÜķw×÷é“ŋšÔĘÕŧ¯Æ÷”ŖŦŧ¯ÖĐÉŲÁŋŋšÔŗĘßfŊoŨoÖúTŧ°ûĄŖŌō´ËŖŦÔÚŗõ´Î¸ĐČžĩÄĪāĻēķÆÚģōÍŦŌģšĨôÕßëSēķ¸ĐČžĩÄÔįÆÚŖŦBŧ°û×÷éŋšÔŗĘßfŧ°û°l]ÖøÖØŌĒ×÷ĶÃĄŖ

- GelNEST™ģųŲ|ÄzÖúÁĻĒĻŲQŌ׹ڞĪÂĩÄŋÆŅвģ´_ļ¨ĐÔ
- ĀÂüŧŧĐgÔÚδíátWÖĐĩÄĒĶÃátš¤ČÚēĪÉŗũŗÉšĻÅeĐĐ
- ˛ŠúōvÔŠ2025Ę״δķ´ŲäNé_ĸŖŦŗŦļāģŨĩČÄãí
- ÖĐĖĐÂÖÛé_WģŨ:ŧ°ûōĖ×Ņb980ÔĒ,ÅāđBģųÕÛŋÛĩČ
- ÖĐĖĐÂÖÛĪŪr´ŲäNŖēŲIŧ°ûžÍËÍÖ§ÔķwzyÔŠēĐ
- ČAÍūÖĐxÔŠēĐÄęÄŠĪŪrģŨíŌuŖŦĻÖĩĪíēÃļY
- ÃĀĩÂÂÍÆŗöĐÂÆˇÖØŊMŧ°ûĪĩĶÃĶÚÉúŽaÎŋËÂĄŅĒĐÍÔŠ
- °ŲÎĐĄûÅßŅŋÄũŧ¯ËØĖŊáČžÁĪ8ÕÛģŨŖŦŲIŧ´ŲĘķË|