細胞死亡的新途徑泛凋亡 (PANopotosis)詳述

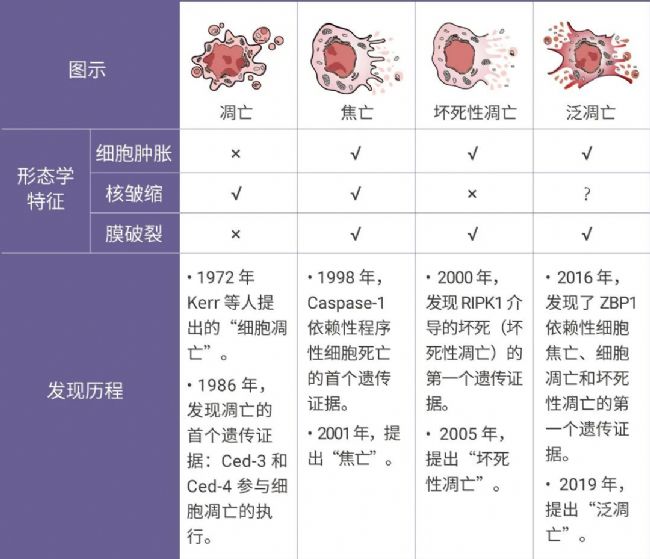
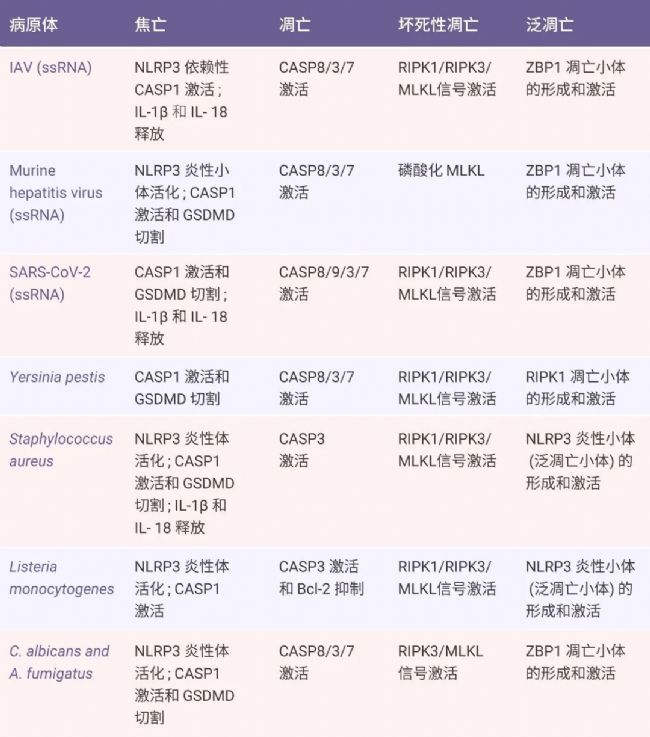
在所有提出的程序性細胞死亡 (PCD) 形式中,細胞焦亡、細胞凋亡和壞死性凋亡是最明確的,其復雜的分子機制負責細胞死亡的啟動、轉導和執行[1]。但實際上這三種途徑之間存在復雜的串擾。例如在無菌損傷和感染性疾病(例如甲型流感病毒 IAV 感染)中,研究人員觀察到所有三種 PCD 途徑的生化標記物均被激活[1]。
2019 年,美國學者 Malireddi 將這種具有焦亡、凋亡和壞死性凋亡特征的新型死亡方式命名為泛凋亡,并提出先天性免疫傳感器 ZBP1 和 TAK1 激酶在泛凋亡小體復合物組裝的調控中發揮重要作用[2]。 Tips:
Tips:泛凋亡(PANoptosis) 是由特定觸發器激活并 PANoptosome 復合物調節的炎癥性 PCD 途徑,其綜合了細胞焦亡、細胞凋亡和/或壞死性凋亡的關鍵特征,這也是 PANoptosis 術語中 “P” “A” 和 “N” 的來源,但其不能被細胞焦亡、凋亡和壞死性凋亡中任意一種死亡方式單獨表征[1]。

多種蛋白質可以形成調節 PCD 的多蛋白復合物,其基于各種蛋白質結構域之間的相互作用,可分為三類:傳感結構域、組裝結構域和催化結構域[2]。
▐ 泛凋亡小體: PANoptosome
泛凋亡受到上游感受器和分子信號的級聯調控,這些傳感器和級聯信號組裝成一種多聚體復合物,即泛凋亡小體 (PANoptosome)。PANoptosome 及其上游的感受器不僅是下游分子的激活平臺,也是 3 種 PCD 途徑的啟動“總開關”[4][5]。
PANoptosome 小體作為一種分子支架,允許參與焦亡、細胞凋亡和/或壞死性凋亡的關鍵分子進行耦聯和結合[1]。傳感器蛋白在感知到病原體成分后,會介導 RIPK3、RIPK1、CASP8、FADD等蛋白組裝成 PANoptosome 復合體,從而誘導 PANoptosis[2][6]。
構成 PANoptosome 小體的蛋白質通常可分為三類:(1) ZBP1 和 NLRP3 作為假定的 PAMP 和 DAMP 傳感器,(2) ASC 和 FADD 作為適配器,以及 (3) RIPK1、RIPK3 、CASP1 和 CASP8 作為催化效應器[2]。
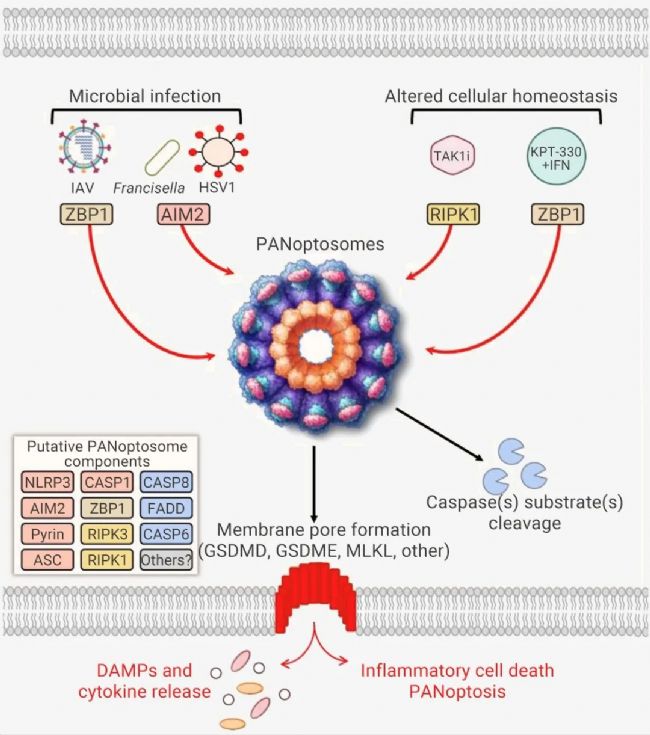
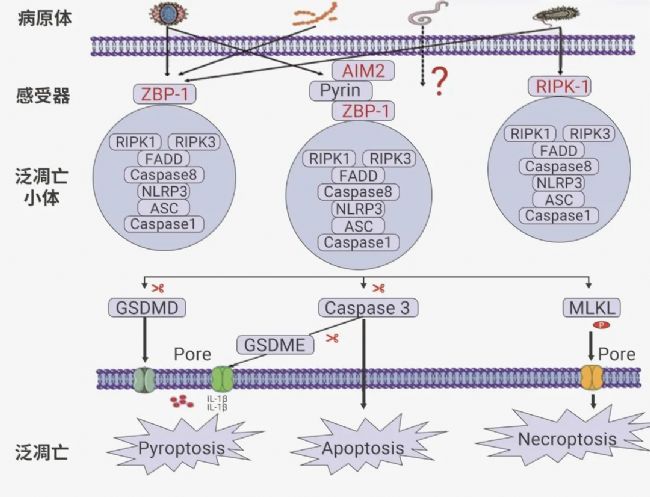
圖 3. 泛凋亡小體及其通路[4]。
 Tips:
Tips:(1) ZBP1 泛凋亡小體:Z-DNA 結合蛋白 1 (ZBP1) 是一種先天免疫受體,被鑒定為甲型流感病毒(Influenza A virus,IAV)的感受器和 IAV 感染期間細胞死亡的主要調節因子。在 IAV 刺激下,ZBP1 作為特異的感受器被激活,進而觸發 PANoptosome 的組裝[7][8]。ZBP1-PANoptosis 形成后,可以啟動由焦亡、凋亡和壞死性凋亡成分組成的泛凋亡,最終導致溶解性炎癥細胞死亡,其特征是 Caspase-1、Caspase-3、Caspase-8 的激活和 MLKL 的磷酸化[9]。
(2) RIPK1 泛凋亡小體:RIPK1 介導的泛凋亡小體主要由 RIPK1、ASC、Caspase-1、Caspase-8 和死亡相關結構域蛋白(fas-associated protein with death domain,FADD)構成,最早由 Malireddi 教授團隊提出。該團隊還發現 TAK1 可作為 RIPK1-PANoptosome 的調控主開關,因為 TAK1 基因的缺失或者功能失活可以觸發 RIPK1-PANoptosome 的組裝。然而,后續研究發現,TAK1 的缺乏同樣可以誘導 RIPK1 非依賴的泛凋亡通路的激活,后者主要由 RIPK3-MLKL 介導。
(3) AIM2 泛凋亡小體:其組成分子包括 AIM2、ZBP1、pyrin、ASC、Caspase-1、Caspase-8、RIPK1、RIPK3 和 FADD[10]。AIM2 炎性小體可感知雙鏈 DNA,AIM2 調節先天免疫傳感器 Pyrin 和 ZBP1 來驅動炎癥信號傳導和炎癥細胞泛凋亡,在人類的正常發育、傳染性疾病、炎癥和腫瘤方面發揮重要作用。

說了這么多,這種 “三合一” 的復雜細胞死亡方式,又該如何研究及檢測的呢?
首先咱們來說檢測,由于泛凋亡綜合了細胞焦亡、細胞凋亡和/或壞死性凋亡的關鍵特征,因此在進行相關檢測時三方面缺一不可!
之前小 M 也為大家整理過相關的檢測方法,那么本期僅做整理,不再詳述。
 Tips:
Tips:
泛凋亡檢測方法及指標!
(1) 觀察細胞形態:細胞焦亡引起細胞質腫脹和膜破裂;凋亡主要形態學特征包括染色質固縮、DNA 片段化、細胞膜起泡、細胞皺縮、凋亡小體的形成等。(2) 檢測不同 PCD 的關鍵蛋白:焦亡相關:Caspase-1、Caspase-3、Gasdermins、AIM2/Pyrin/NLRP3 等;凋亡相關:Caspase-3、Caspase-7、Caspase-8、PARP、Bax/Bcl 等;壞死性凋亡相關:MLKL、RIPK1、RIPK3、ZBP1 等。
(3) 其他指標檢測:Annexin V-FITC 和 PI 聯合染色;TUNEL 法;JC-1 檢測;ELISA 檢測炎癥因子的釋放;免疫印跡、流式等技術檢測炎性小體 NLRP3 的表達和 Caspase-1 的活化情況。
▐ 文獻案例解讀(IF=39.3):
近期,Jin-Fei Lin 等人發現磷酸化的 NFS1 可通過預防 PANoptosis 以削弱結直腸癌的 Oxaliplatin 敏感性[11]。
作者使用基于體內代謝酶基因的 (CRISPR)-Cas9 文庫篩選,發現 NFS1 的缺失顯著增強了 CRC 細胞對奧沙利鉑的敏感性。體外和體內結果表明,NFS1 缺陷與 Oxaliplatin 協同作用,通過增加細胞內活性氧 (ROS) 水平,引發泛凋亡。
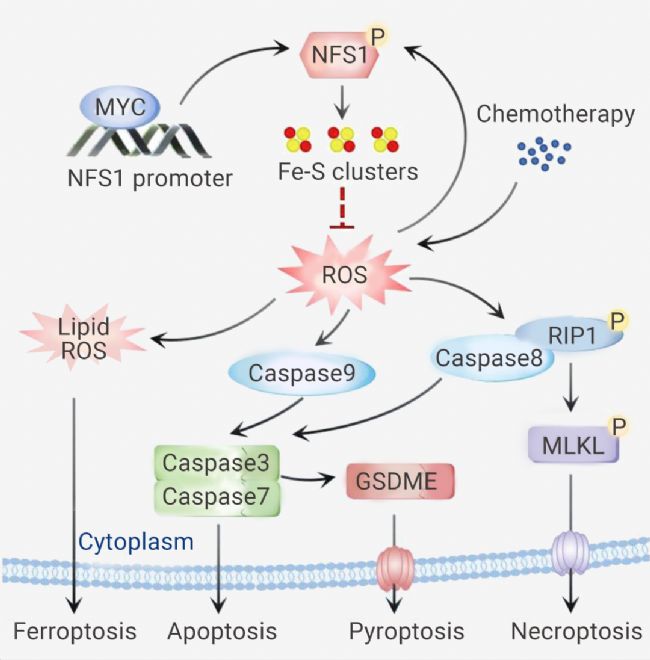
圖 4. 磷酸化的 NFS1 通過降低 ROS 水平來減弱鉑基化學敏感性,從而防止 PANoptosis[11]。
為了研究發生的細胞死亡類型,使用常見細胞死亡途徑的多種抑制劑并結合光鏡觀察細胞形態、YP1/PI 染色、流式檢測細胞死亡等進行檢測。結果表明,焦亡 (GSDME) 抑制劑 Disulfiram 和自噬抑制劑 3-methyladenine 無顯著作用,而凋亡抑制劑 Z-VAD-FMK 、壞死性凋亡抑制劑 Necrostatin-1 、鐵死亡抑制劑 Ferrostatin-1 、焦亡 (GSDME) 抑制劑 Ac-DMPD/DMLD-CMK 逆轉(但不能完全恢復)Oxaliplatin 治療下由 NFS1 缺乏引起的細胞活力降低和細胞毒性增強 (圖 5)。
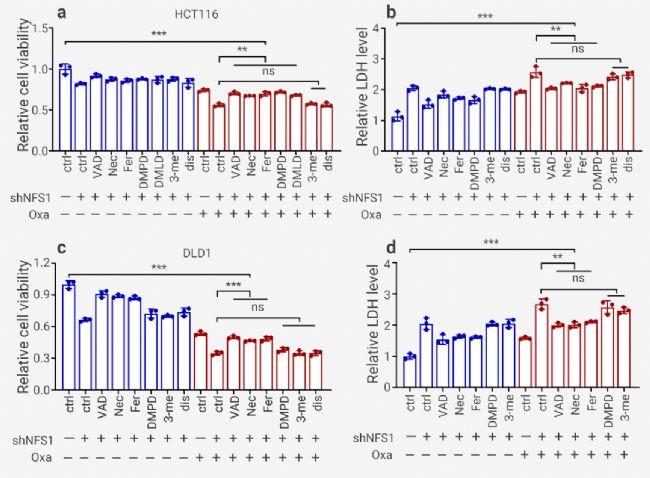
圖 5. Oxaliplatin 聯合多種抑制劑的細胞活力及毒性測定[11]。
a-b: 對照和 NFS1 敲低 HCT116 細胞的細胞活力 (a) 和細胞毒性 (b) 評估; c-d: 對照和 NFS1 敲低 DLD1 細胞活力 (c) 和細胞毒性 (d) 評估。
為了進一步證實 PANoptosis 的發生,作者發現 NFS1 敲低聯合 Oxaliplatin 治療顯著增加了死亡細胞的數量,包括指示細胞凋亡或壞死性凋亡的 YP1 陽性細胞和指示細胞壞死性凋亡、細胞焦亡或鐵死亡的 PI 陽性細胞 (圖 6: 左圖, a-b 和右圖 a-b)。
對 HCT116 細胞的分析還表明,聯合組中從質膜出現的大氣泡數量明顯高于陰性對照組、NFS1 敲低組和 Oxaliplatin 治療組,這表明細胞焦亡的發生 (圖 6: 左圖, a)。此外,NFS1 耗竭后進行 Oxaliplatin 治療定量增加了早期/晚期凋亡細胞的數量,并顯著減少了活細胞的數量 (圖 6: 左圖, c-d 和右圖 c-d)。此外,在 NFS1 敲低組和 Oxaliplatin 治療組中檢測到的脂質 ROS 含量高于對照組。這種增加在聯合組中更為明顯,表明鐵死亡的發生 (圖 6: 左圖, e 和右圖 e-f)。因此,這些數據表明 NFS1 缺陷與 Oxaliplatin 聯合有助于 PANoptosis 的激活。
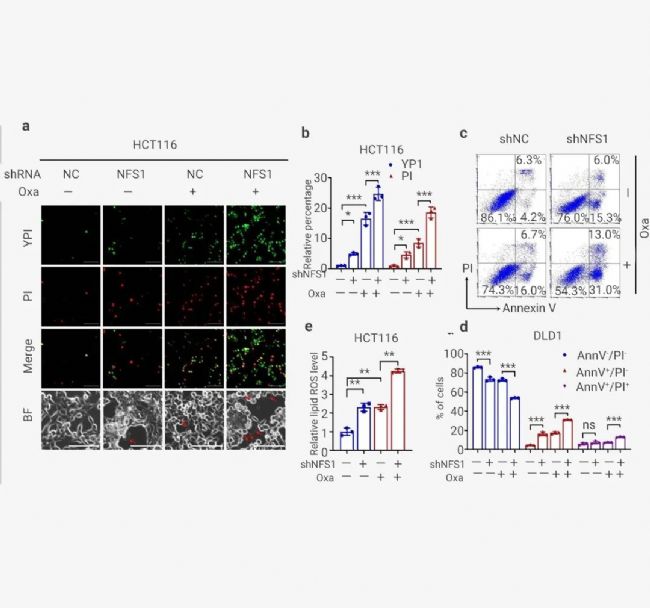
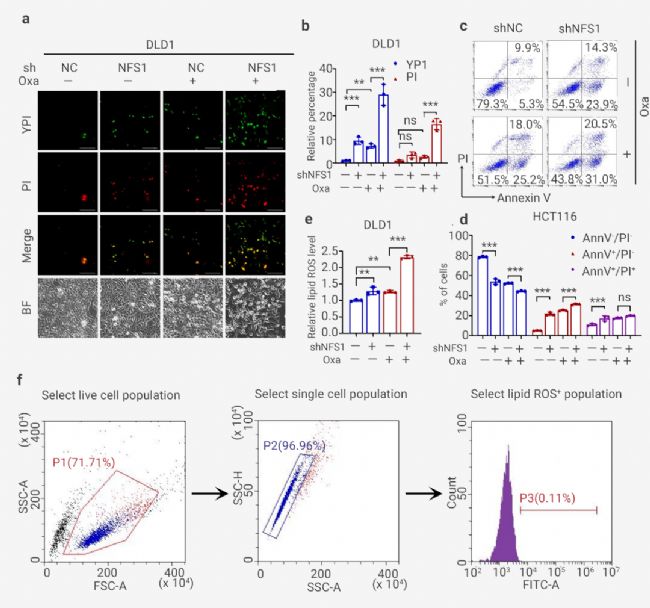 圖 6. NFS1 缺陷與 Oxaliplatin 治療協同作用,誘導 PANoptosis[11]。
圖 6. NFS1 缺陷與 Oxaliplatin 治療協同作用,誘導 PANoptosis[11]。 (左圖): a. 對照中可能發生凋亡或壞死性凋亡的 YP1+細胞(綠色)和可能發生凋亡、壞死性凋亡、細胞焦亡或鐵死亡的 PI+細胞(紅色)以及用 Oxaliplatin 處理的 NFS1 敲低 HCT116 細胞。底部顯示為明場,紅色箭頭表示從質膜中出現的大氣泡; b. 對 (a) 中的 YP1+ 和 PI+ 細胞進行定量。c-d: 流式細胞術 (c) 和采用膜聯蛋白 V/PI 染色的定量分析 (d),在對照和用 PBS 或 Oxaliplatin 處理的 NFS1 敲低 DLD1 細胞中評估活細胞 (Annexin V- /PI-)、早期凋亡細胞 (Annexin V+ /PI-) 和晚期凋亡細胞的百分比 (Annexin V+ /PI+); e. 通過 BODIPY™ 581/591 C11 探針測定評估用 Oxaliplatin 處理的對照和 NFS1 敲低 HCT116 細胞中的脂質 ROS 水平。
(右圖): a. 對照中可能發生凋亡或壞死性凋亡的 YP1+細胞(綠色)和可能發生凋亡、壞死性凋亡、細胞焦亡或鐵死亡的 PI+ 細胞(紅色)以及用 Oxaliplatin 處理的 NFS1 敲低 DLD1 細胞。底部顯示為明場; b. 對 (a) 中的 YP1+ 和 PI+ 細胞進行定量; c. 使用膜聯蛋白 V/PI 染色進行流式細胞術分析,評估對照和用 PBS 或 Oxaliplatin 處理的和 NFS1 敲除 HCT116 中活細胞 (Annexin V-/PI-)、早期凋亡細胞 (Annexin V+/PI-) 和晚期凋亡細胞 (Annexin V+/PI+) 的百分比; d. (c)中活細胞 (Annexin V-/PI-)、早期凋亡細胞 (Annexin V+/PI-) 和晚期凋亡細胞 (Annexin V+/PI+) 的定量; e. 使用 BODIPYTM 581/591 C11 探針測定法評估Oxaliplati 處理的對照細胞和 NFS1 敲低 DLD1 細胞中的脂質 ROS; F. 本研究中用于 ROS/脂質 ROS 分析的策略。
此外,作者評估了 Oxaliplatin 治療下 NFS1 缺陷誘導 PANoptosis 的具體機制。總之,Oxaliplatin 介導的氧化應激可增強 NFS1 的絲氨酸磷酸化水平,并且 NFS1 以 S293 磷酸化依賴性方式阻止 Oxaliplatin 治療下 PANoptosis 的激活。
 本期小 M 為大家介紹了細胞死亡的新姿勢:泛凋亡 (PANopotosis),一種炎癥性細胞程序死亡途徑。三種重要的泛凋亡小體 ZBP1-PANoptosomes、AIM2-PANoptosomes 和 RIPK1-PANoptosomes 在其中發揮重要的調節作用,此外,小 M 為大家介紹了泛凋亡的檢測指標,并通過文獻案例進一步詳述。希望大家能夠有所收獲喔~
本期小 M 為大家介紹了細胞死亡的新姿勢:泛凋亡 (PANopotosis),一種炎癥性細胞程序死亡途徑。三種重要的泛凋亡小體 ZBP1-PANoptosomes、AIM2-PANoptosomes 和 RIPK1-PANoptosomes 在其中發揮重要的調節作用,此外,小 M 為大家介紹了泛凋亡的檢測指標,并通過文獻案例進一步詳述。希望大家能夠有所收獲喔~

|
Oxaliplatin 是一種 DNA 合成 抑制劑。Oxaliplatin 會導致 DNA 交聯損傷,阻止 DNA 復制和轉錄并導致細胞死亡。Oxaliplatin 以時間依賴方式抑制人黑色素瘤細胞系 C32 和 G361,IC50 值分別為 0.98 μM 和 0.14 μM。Oxaliplatin 可以誘導細胞自噬 (autophagy)。 |
|
Disulfiram (Tetraethylthiuram disulfide) 是特異性的 ALDH1 抑制劑,對酒精具有急性敏感性。Disulfiram 有效抑制人體和小鼠細胞中脂質體中的 GSDMD 孔形成,炎性體介導的細胞凋亡和 IL-1β 分泌。Disulfiram + Cu2+ 可增加細胞內 ROS 水平,引發卵巢癌干細胞凋亡。 |
|
Daurisoline 是一個 hERG 抑制劑,也是一種自噬抑制劑。 |
|
Z-VAD-FMK (Z-VAD(OH)-FMK) 是一種 pan caspase 抑制劑。Z-VAD-FMK 不抑制泛素 C 末端水解酶 L1 (UCHL1) 活性,即使濃度高達 440 μM。 |
|
Necrostatin-1 (Nec-1) 是一種有效的能透過血腦屏障的壞死性凋亡 (necroptosis) 抑制劑,在 Jurkat 細胞中的 EC50 為 490 nM。Necrostatin-1 抑制 RIP1 激酶 (EC50=182 nM) 。Necrostatin-1 (Nec-1) 也是一種 (IDO) 抑制劑。 |
參考文獻:
[1] Wang Y, et al. From pyroptosis, apoptosis and necroptosis to PANoptosis: A mechanistic compendium of programmed cell death pathways. Comput Struct Biotechnol J. 2021 Aug 3;19:4641-4657.
[2] Malireddi RKS, et al. ZBP1 and TAK1: Master Regulators of NLRP3 Inflammasome/Pyroptosis, Apoptosis, and Necroptosis (PAN-optosis). Front Cell Infect Microbiol. 2019 Nov 26;9:406.
[3] Zhu P, et al. Advances in mechanism and regulation of PANoptosis: Prospects in disease treatment. Front Immunol. 2023 Feb 9;14:1120034.
[4] Pengyue Zhao, et al. Research progress on the role of PANoptosis in human diseases. August 2022Zhonghua Yi Xue Za Zhi 102(32):2549-2554.
[5] Christgen S, et al. Identification of the PANoptosome: A Molecular Platform Triggering Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front Cell Infect Microbiol. 2020 May 29;10:237.
[6] Samir P, et al. The PANoptosome: A Deadly Protein Complex Driving Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front Cell Infect Microbiol. 2020 Jun 3;10:238.
[7] Gullett JM, et al. It's All in the PAN: Crosstalk, Plasticity, Redundancies, Switches, and Interconnectedness Encompassed by PANoptosis Underlying the Totality of Cell Death-Associated Biological Effects. Cells. 2022 Apr 29;11(9):1495.
[8] Zheng M, et al. Newly Identified Function of Caspase-6 in ZBP1-mediated Innate Immune Responses, NLRP3 Inflammasome Activation, PANoptosis, and Host Defense. J Cell Immunol. 2020;2(6):341-347.
[9] Zheng M, et al. The regulation of the ZBP1-NLRP3 inflammasome and its implications in pyroptosis, apoptosis, and necroptosis (PANoptosis). Immunol Rev. 2020 Sep;297(1):26-38.
[10] Lee S, et al. AIM2 forms a complex with pyrin and ZBP1 to drive PANoptosis and host defence. Nature. 2021 Sep;597(7876):415-419.
[11] Lin JF, et al. Phosphorylated NFS1 weakens oxaliplatin-based chemosensitivity of colorectal cancer by preventing PANoptosis. Signal Transduct Target Ther. 2022 Feb 28;7(1):54.