´úÖxŊMWÔÚŅĐžŋĄ°žÁŖķwéW qĄąŊŌĘžŧ°ûĄ°ˇĩĀĪßͯĄąĩÄĐÂWÃØÖĐĩÄĒĶÃ
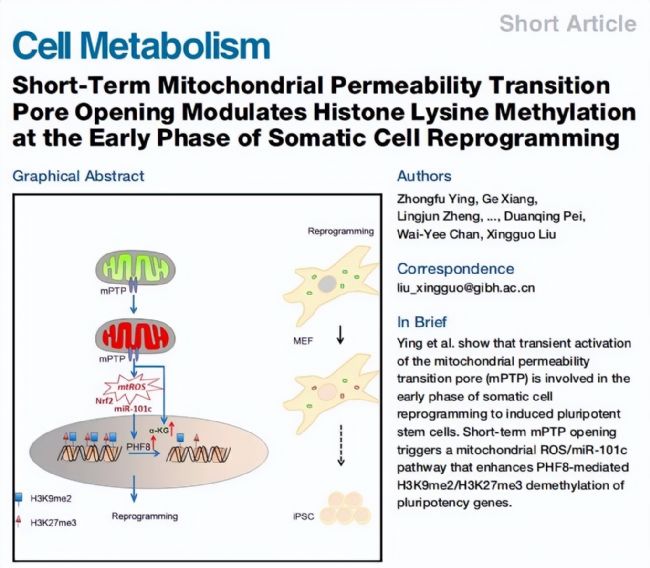
°ląíÆÚŋ¯ŖēCell Metabolism
Ķ°íŌō×ĶŖē22.415
°ląírégŖē2018Äę
ēĪ×÷ÎÎģŖēÖĐøŋÆWÔēVÖŨÉúÎīátËÅcŊĄŋĩŅĐžŋÔē
ŊņĖėBIOTREE°ŲȤ´úÖxŊMWĸŊo´ķŧŌˇÖĪí°ĸȤÉúÎī
fÖúŋÍô°ląíÔÚCell MetabolismÉĪĩÄŌģÆĒÎÄÕÂŖēShort-Term Mitochondrial Permeability Transition Pore Opening Modulates Histone Lysine Methylation at the Early Phase of Somatic Cell ReprogrammingĄŖ
Ę˛Ã´ĘĮ“žÁŖķwéW q”?MŋįžÁŖķwČÍâĤĩÄžÁŖķwͨ͸ĐÔŪDQŋ×Ŗ¨mPTPŖŠĩÄ˲ré_ˇÅˇQé“žÁŖķwéW q”[1]ĄŖmPTPÔÚŧ°ûĩÄÉú´æēÍĩōÍöÖĐÆđĩŊÁËQļ¨ĐÔĩÄŌâÁxĄŖŋ×ÍęČĢé_ˇÅū§ÖÂŧ°ûĩÄĩōÍöŖŦŋ×ĩÄ˲ré_ˇÅūÕ{ŋØŧ°ûĩÄ°lĶũĄŖ
°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦžÁŖķwÔÚļāÄܸÉŧ°ûŖ¨ŌģˇNžßĶĐ×ÔÎŌ¸üĐÂŖŦ×ÔÎŌÍÖÆÄÜÁĻĩÄļāÄÜŧ°ûŖŠĩÄ°lĶũÖĐÆđĩŊÖØŌĒ×÷ĶÃŖŦļøķwŧ°ûÔŲžŗĖĩŊÕT§ļāšĻÄܸÉŧ°ûÖØČžÉĢŲ|ĩÄĐŪīŖŦß@Ōģß^ŗĖĘĮˇņŌÔŧ°ČįēÎͨß^žÁŖķwÖĐĩÄĐÅĖßMĐĐÕ{šß˛ģĩÃļøÖĒĄŖ
ÄĮÃ´Ę˛Ã´ĶÖĘĮķwŧ°ûÖØžŗĖÄØŖŋĻŌģˇÖģ¯ŗÉĘėĩÄŧ°ûíÕfŖŦÆäŧ°ûēËČĢÄÜĐÔĩÄŦFĘĮŊ¨ÁĸÔÚÅcÂŅŧ°ûŲ|ČÚēĪģųĩAÉĪĩÄĄŖß@ˇNĶÉķwŧ°ûĪōČĢÄÜŧ°ûĩÄŪDסQéķwŧ°ûÖØžŗĖ[2]ĄŖ´ķŧŌĘėÖĒĩÄŋËÂĄŅōļāĀōžÍĘĮß@ô“ĘĀ”ĩÄĄŖ
ĩĢŦFÔÚŌĒŦFß@ŌģÄŋĩÄŖŦķwŧ°ûēËŌÆÖ˛˛ĸ˛ģĘĮΨŌģĘÖļÎĄŖ2006ÄęČÕąžŋÆWŧŌÉŊÖĐÉė(Shinya Yamanaka)Ę×´ÎĀûĶÃ˛ĄļžŨdķwĸËÄŪDäŌō×ĶĩÄŊMēĪŪDČëˇÖģ¯ĩÄķwŧ°ûÖĐŖŦĘšÆäÖØžŗĖļøĩÃĩŊÁËîËÆÅßĖĨ¸Éŧ°ûĩÄŌģˇNŧ°ûîĐÍ——ÕT§ļāÄܸÉŧ°û(iPSCs)ĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦÔŗÉšûÔÚ2012ÄęąģĘÚĶčÁËÖZØ ÉúĀíW/átWĒĄŖ
mPTPĩÄé_ˇÅŧ°ęPé]Ļŧ°ûĩÄĩōÍöŧ°°lĶũŽaÉúĩÄĶ°íß@ąŗēķĘĮˇņŌÔŧ°ĘĮČįēÎÕ{ŋØŧ°ûēËĩÄßĘĮδÖĒĩÄĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦģųĶÚŌÔÉĪŊYšûŖŦWÕßĸŅĐžŋĩÄÖØĐÄŪDŌÆÖÁmPTPĄŖŅĐžŋ°lŦFÔÚÖØžŗĖĩÄÔįÆÚmPTPŊvÁËļĖÆÚé_ˇÅŖŦß@ˇNļĖēĩÄŧ¤ģîÔöÁËÖØžŗĖĄŖļĖÆÚmPTPĩÄ´ōé_Ķ|°lžÁŖķwROS/miR-101cͨ¡ŖŦÔöPHF8Ŗ¨ŊYÍŦÔ´Ö˛ÎīĶōÖ¸ĩ°°×ŖŠŊé§ĩÄļāÄÜĐÔģųŌōĩÄH3K9me2/H3K27me3ČĨŧ×ģųģ¯ŖŦŊĩĩÍÆäÔÚļāÄÜģųŌōĸĶ×Ķ
^ĩÄÕŧĶĐÂĘŖŦĖá¸ßÖØžŗĖЧÂĘĄŖ
ÔÚÖØĐžŗĖĩÄÔįÆÚëAļÎ
mPTPĩÄļĖÆÚé_ˇÅ
éÁË´_ļ¨mPTPÔÚÖØžŗĖß^ŗĖÖĐĩÄ îBŖŦĻĐĄĘķÅßĖĨŗÉĀwžSŧ°û(MEFs)ÔÚSox2ĄĸKlf4ĄĸOct4Ąĸc-Myc (SKOM)Ŋé§ĩÄÖØžŗĖß^ŗĖÖĐĩÄmPTPßMĐĐÁËąOyĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦŅĐžŋ°lŦFmPTPĩÄé_ˇÅļČÔÚĩÚ3ĖėÔöŧĶŖŦÔÚĩÚ5ĖėÔŲ´ÎĪÂŊĩŖŦ˛ĸÔÚĩÚ8ĖėąŖŗÖß@ËŽÆŊŖŦâ}üSžGËØÉšâČžŠŖ¨CalceinŖŠÅcmPTPé_ˇÅŗĖļČŗĘØĪāęPŖ¨DA, BŖŠĄŖ
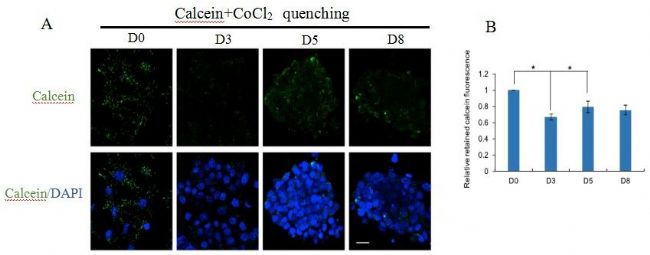
×ĸŖēA: š˛žÛŊšDĪņÔÚÖØĐžŗĖĩÄĩÚ0Ąĸ3Ąĸ5ēÍ8ĖėˇÖÎömPTPĩÄé_ˇÅĮérĄŖĶÆˇÅccalceinēÍCoCl2š˛ŨdĄŖB: ŧ°ûĪāĻÁôâ}üSžGËØÉšâÔÚÖØžŗĖß^ŗĖÖСÖeĶÚĩÚ0Ąĸ3Ąĸ5Ąĸ8ĖėĩÄļ¨ÁŋĮérĄŖ
ÔįÆÚĩÄmPTPé_ˇÅÔöÁËÖØĐžŗĖ
éÁË´_ļ¨mPTPÔÚÖØžŗĖÖĐÆđĩŊĩÄ×÷ĶÃŖŦŅĐžŋČËT
^ˇÖÁË°üēŦŪDģųŌō Oct4 ´ŲßMŌōËØōĶžGÉĢÉšâĩ°°×Ŗ¨GFPŖŠąíß_ÔÚČĩÄMEFs (ĐĄĘķÅßĖĨŗÉĀwžSŧ°û)ŖŦ¸ųūâ}üSžGËØÉšâĩÄļČ´_ļ¨mPTPĩġÅé_ĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦDÖĐŋÉŌÔŋ´ŗö¸ßmPTPé_ˇÅŧ°ûĩÄÖØžŗĖЧÂʸßĶÚĩÍmPTPé_ˇÅĩÄЧÂĘŖ¨DCŖŠĄŖ
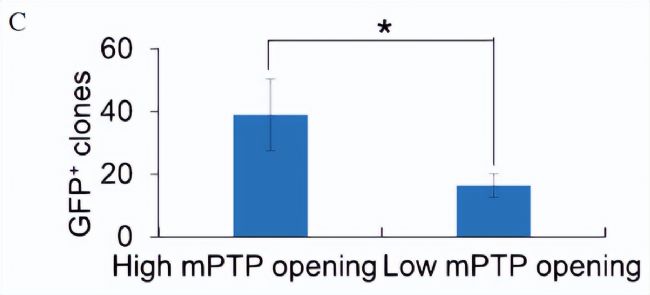
×ĸŖēĻ¸ßĄĸĩÍmPTPé_ˇÅļČĩÄskom¸ĐČžŧ°ûßMĐĐÅÅĐōēķŖŦļ¨ÁŋˇÖÎöÖØŊMЧÂĘĄŖ
éÁËßMŌģ˛ŊŅĐžŋmPTPÔÚÖØžŗĖÖĐĩÄ×÷ĶÃŖŦŅĐžŋČËTͨß^ÔöŧĶģōpÉŲšĻÄÜĩġŊˇ¨íÕ{šmPTPĩÄ´ōé_ĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦËû°lŦFęPæIĩÄmPTPŊMˇÖĶHhËØDŖ¨CypDŖŠĩÄß^ļČąíß_§ÖÂmPTP´ōé_ŖŦļøCypDŗÁÄŦŌÖÖÆmPTP´ōé_ŖŦß@ĘĮŌģˇNžÁŖķwģųŲ|ëÄģų¸Ŧ°ąõŖíˇ´ĘŊŽÃ¸ŖŦÄŋĮ°ŌŅÖĒĩÄŋÉÕ{šmPTPĩÄé_ˇÅ(DD)ĄŖ
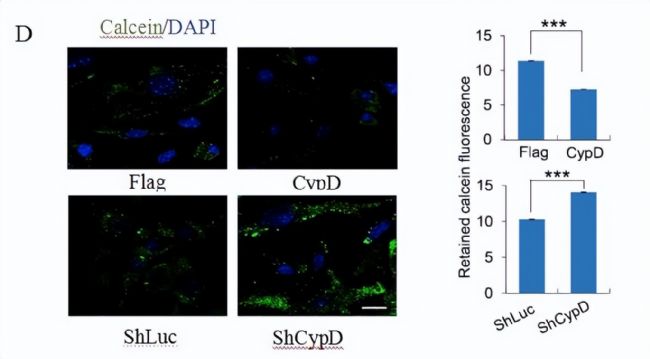
×ĸŖēD: ÔÚCypDß^ąíß_ģōŗÁÄŦĩÄlŧūĪÂŖŦĶÃshRNAˇÖÎömPTPĩÄé_ˇÅĮérĄŖ×ķDéĩäĐÍĩÄš˛ŊšDĪņŖŦĶŌDéÁôâ}üSžGËØÉšâĩÄļ¨ÁŋĄŖ
CypDß^ąíß_H3K9me2ēÍH3K27me3
ĩÄČĨŧ×ģųģ¯ŌĀŲĶÚPHF8
zyÁËĶĐģō]ĶĐCypDß^ąíß_ĩÄŧ°ûÖØžŗĖĩÚ3ĖėŊMĩ°°×ĩÄŧ×ģųģ¯ îBĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦō°lŦFŖŦÅcĻÕÕŊMĪāąČŖŦÔÚCypDß^ąíß_ÖØžŗĖĩÄĩÚ3ĖėēÍĩÚ4ĖėŖŦH3K9me2ēÍH3K27me3ĩÄËŽÆŊĪÂŊĩ(DE)ŖŦļøÆäËûyÔŊM]ĶĐī@ĘžÃ÷ī@ĩÄ×ģ¯ĄŖͨß^H3K9me2ēÍH3K27me3ČžÉĢŲ|ÃâŌßŗÁĩí(ChIP)ōˇÖÎöÆäÕŧĶÃÂĘŖŦ°lŦFCypDß^ąíß_ŊĩĩÍÁËÆäÔÚSox2ĄĸNanogĄĸOct4ĩČļāÄÜģųŌōĸĶ×Ķ
^ĶōĩÄÕŧĶÃÂĘĄŖ
Ōō´ËŖŦmPTPĩÄŧ¤ģî§ÖÂH3K9me2ēÍH3K27me3ĩÄÕûķwČĨŧ×ģųģ¯ĄŖH3K9ēÍH3K27ĩÄŧ×ģųģ¯ĘÜļāˇNŊMĩ°°×ČĨŧ×ģųģ¯Ã¸ĩÄÕ{ŋØŖŦéÁËÕŌŗöÄÄĐŠŊMĩ°°×ČĨŧ×ģųģ¯Ã¸
ĸÅcÁËmPTPĩÄé_ˇÅĄŖÎŌzyÁ˲ģÍŦŊMĩ°°×ČĨŧ×ģųģ¯Ã¸ÔÚĶĐģōoCypDß^ąíß_ĩÄŧ°ûSKOMÖØžŗĖß^ŗĖÖĐĩÄąíß_ËŽÆŊĄŖŊYšûąíÃ÷ŖŦÔÚąģzyĩÄĩ°°×ÖĐŖŦÖģĶĐPHF8ÔÚCypDß^ąíß_rÔöŧĶ(DF)ĄŖ´ËÍâŖŦŅĐžŋČËTÔÚÖØžŗĖÖĐĘšĶÃshRNAĻPHF8ßMĐĐŗÁÄŦŖŦ°lŦFĮÃŗũPHF8ŋÉŌÔģÖÍH3K9me2ēÍH3K27me3ĩÄËŽÆŊŖŦ˛ĸͨß^CypDß^ąíß_Ī÷ČõÖØžŗĖЧÂĘĩÄĖá¸ß(DG)ĄŖ
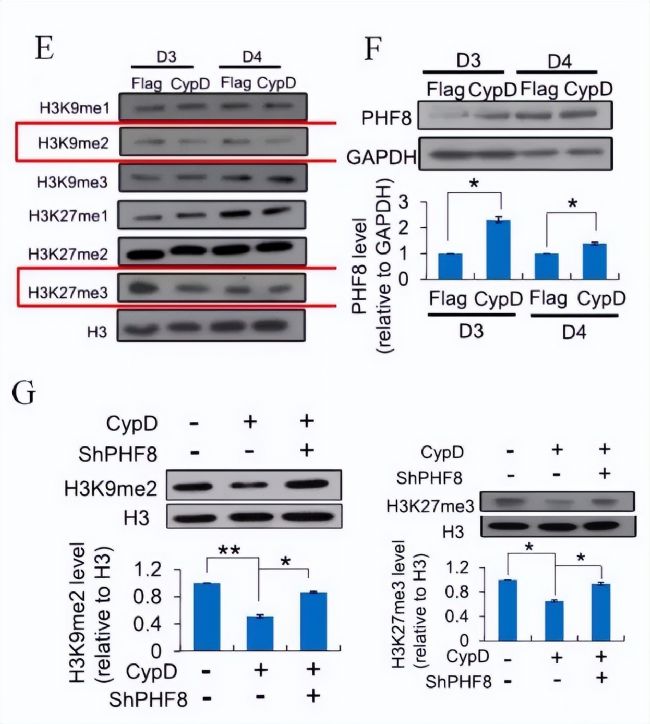
×ĸŖēE: ÖØŊMß^ŗĖÖĐCypDß^ąíß_ĩÚ3ĖėēÍĩÚ4ĖėēķH3K9me1/2/3ēÍH3K27me1/2/3ĩÄWestern blotˇÖÎöĄŖF: ÖØŊMēķĩÚ3ĖėēÍĩÚ4ĖėCypDß^ąíß_ŧ°ûÖĐPHF8ĩÄWestern blotˇÖÎöĄŖG: H3K9me2ēÍH3K27me3ŊPHF8 shRNAĖĀíēͲģŊPHF8 shRNAĖĀíĩÄCypDß^ąíß_ŧ°ûÔÚÖØžŗĖēķĩÚ3ĖėĩÄWestern blotˇÖÎöĄŖ
°ĐË´úÖxŊMWĻα-KGßMĐĐļ¨ÁŋˇÖÎö
PHF8´ßģ¯Ų°ąËáČĨŧ×ģųģ¯ŖŦĐčŌĒa-ÍĒÎėļūËá(a- KG)ŖŦß@ĘĮČũôČËáŅhĩÄÖĐég´úÖxŽaÎīŖŦ˛ÉĶÃŗŦ¸ßЧŌēĪāÉĢ×V´ŽÂŲ|×Vˇ¨(UHPLC-MS/MS)ßMĐĐļ¨Áŋyļ¨´úÖxÎīËŽÆŊŖŦŅĐžŋČËTͨß^´úÖxÎīˇÖÎöíļ¨ÁŋÖØžŗĖÖĐCypDß^ąíß_ēķĩÄa-KGËŽÆŊĄŖ°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦCypDß^ąíß_ŧ°ûĩÄa-KGËŽÆŊ¸ßĶÚĻÕÕŊM(DH)ĄŖŌō´ËŖŦa- KGĩÄÔöŧĶŋÉÄÜĘĮPHF8ģîĐÔÔöŧĶĩÄŌģŌōËØĄŖ
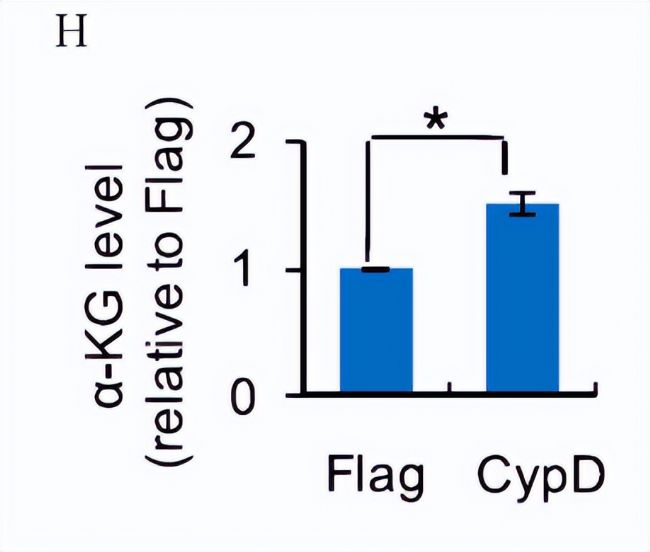
×ĸŖēÖØŊMēķĩÚ3ĖėFlagēÍCypDß^ąíß_ŧ°ûÖĐa-KGËŽÆŊĩĸßЧŌēĪāÉĢ×V-Ų|×VˇÖÎö
ŊYÕZ
ąžō°lŦFžÁŖķwͨ͸ĐÔŪDQŋ×Ŗ¨mPTPŖŠÔÚÖØžŗĖĩÄÔįÆÚŊvÁËļĖÆÚé_ˇÅŖŦmPTPĩÄé_ˇÅĘšH3K9me2ēÍH3K27me3°lÉúČĨŧ×ģųģ¯ŖŦ§ÖÂËüÔÚŧ°ûÖĐĩÄļāÄÜĐÔģųŌōĩÄĸĶ×Ķ ^ĶōÕŧĶĐÂĘŊĩĩÍĄŖmPTPĩÄé_ˇÅÔöŧĶÁËžÁŖķwģîĐÔŅõŽaÎīĪÂĶÎĩÄPHF8ĩ°°×ËŽÆŊēÍmir-101cŖŦÍŦrŌ˛Ėá¸ßÁËPHF8ĩÄŨoŌō×Ķa-ÍĒÎėļūËáĩÄËŽÆŊĄŖÄļø´ŲßMķwŧ°ûŪD×éļāÄܸÉŧ°ûĄŖ
°ŲȤ´úÖxŊMWÎÄĢIˇÖĪíŖŦŅĐžŋŊYšûąíÃ÷ŖŦÔÚmPTPŊé§ĩÄąíĶ^ßz÷Õ{ŋØĪÂŖŦžÁŖķwĩŊēËĩÄĐÂÍžŊQļ¨ÁËŧ°ûĩÄÃüß\ĄŖÍŦrßMŌģ˛ŊĀŊüÁ˸Éŧ°ûēÍÅR´˛ŧ˛˛ĄÖίĩÄžāëxĄŖļøÔˇŊˇ¨ÅcŊĩäĩÄÅßĖĨ¸Éŧ°ûēÍķwŧ°ûēËŌÆÖ˛ŧŧĐg˛ģÍŦĩÄĘĮŖŦËü˛ģĘšĶÃÅßĖĨŧ°ûģōÂŅŧ°ûŖŦËųŌÔ]ĶĐĀíWî}ĄŖëmČģÔˇŊˇ¨ĩÄ°˛ČĢĐÔßĶĐ´ũŋŧÁŋŖŦĩĢÔÚ°l˛ĄCĀíŌÔŧ°ŧ˛˛ĄÔ\āĩČátWˇŊÃæļŧĶĐžŪ´ķĩÄÁĻŖŦÖĩĩÃδíÉîČëĩØŅĐžŋŧ°WÁĄŖ
ĸŋŧÎÄĢI
[1]Ying Z , Chen K , Zheng L , et al. Transient Activation of Mitoflashes Modulates Nanog at the Early Phase of Somatic Cell Reprogramming[J]. Cell Metabolism, 2015, 23(1).
[2]°˛ÍūŖŽátWŧ°ûÉúÎīWŖ¨ĩÚČũ°æŖŠŖēąąžŠ´ķWátWŗö°æÉįŖŦ2013

- Target-BSŧŧĐgŊŌĘžĖĮÄō˛ĄŌũ°l˛ĒÆđšĻÄÜÕĪĩKĩÄDNAŧ×ģųģ¯Õ{ŋØCÖÆ
- ēĪŗÉÉúÎīWĩ׹Pŧ°ûÖŽŽ ŗāŊÍĸŗŖĶÃąíß_Ũdķwŧ°ģųŌō¸ÄÔėŧŧĐg
- ÉîļČŊâÎöCRISPRÎÄėēYßxÁ÷ŗĖŧ°ĒĶð¸Āũ
- ŋÉØēÍlŧūĪ¸ߡֹæÂĘzym6AĐŪīĩÄĐÂm6AyĐōˇŊˇ¨é_°lŅĐžŋ
- AIČËš¤ÖĮÄÜÔÚĩ°°×Ų|ŊYîAyĄĸšĻÄÜîAyŧ°ÔOĶÖĐĩÄ×÷ĶÃ
- m5C MeRIP-seqĩČŊŌĘžm5CĐŪīÔÚ°Š°YÄÍËÖĐĩÄęPæIÕ{ŋØCÖÆÖĐĩÄĒĶÃ
- ChIPŧŧĐgŊŌĘžNURR1ÔÚĮ°ÁĐĪŲ°ŠÄģųŌōŪDäĩŊÄ[ÁößMÕšÖĐĩÄÕ{ŋØCÖÆ
- ɡNŪDäŌō×ĶÔÚŊ駡ŦĮŅšûŗÉĘėĩÄąíĶ^ßz÷Õ{ŋØÖĐ°l]ĩÄęPæI×÷ĶÃ
- 2025˛ŽēĀÉúÎī´ēŧžČĢøŅ˛Övé_ĸŖŦÔįøBķÃûÕŧĪČC
- ČA´ķÖĮÔė¸ąŋ˛ÃÖĐø ^ŋŊĀíÅíggŌģĐĐĩŊÔL˛ŽēĀÉúÎī
- ˛ŽēĀļĘÂéLŗöĪ¯ŖžĢĖØĐÂÖĐĐĄÆķI°lÕšŅ´ķÕ¯˛ĸ°lŅÔ
- ˛ŽēĀÉúÎīëp11ŋņgĖØģŨģîĶíŌuŖŦĪŪrĪŪÁŋ
- âųÃĀͨĩÂÍÆŗöŗÉąžrķwōÎŧ°ûyĐōŖ¨FFPEģōČĢŅĒŖŠ
- ˛ŽēĀÉúÎī2024ĪĩÁĐÅāĶ°āŖ¨ęÎ÷ÕžŖŠķÃûé_ĸ
- ˛ŽēĀÎŧ°ûFlexÄęÖĐžŪģŨŖŦ1.1W/ĶąžģîĶrŧ´ĸŊØÖš
- Ë{ž°ŋÆĐÅDAP-seqŧŧĐgĪāęPÎÄÕÂ6ßB°lŖŦŋIF 95.2